
IMPLEMENTING AN ARTIFICIAL CENTIPEDE CPG
Integrating Appendicular and Axial Movements of the Scolopendromorph
Centipede
Rodrigo R. Braga
Departament of Electronic and Computer Engineering, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
Zhijun Yang
School of Engineering and Electronics, Edinburgh University, Edinburgh EH9 3JL, UK
Felipe M. G. França
Systems Engineering and Computer Science Program, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
Keywords: Centipede locomotion, central pattern generator, distributed algorithms, post-inhibitory rebound, scheduling
by multiple edge reversal.
Abstract: In nature, a high number of species seems to have purely inhibitory neuronal networks called Central
Pattern Generators (CPGs), allowing them to produce biological rhythmic patterns in the absence of any
external input. It is believed that one of the mechanisms behind CPGs functioning is the Post-Inhibitory
Rebound (PIR) effect. Based in the similarity between the PIR functioning and the Scheduled by Multiple
Edge Reversal (SMER) distributed synchronizer algorithm, a generalized architecture for the construction of
artificial CPGs was proposed. In this work, this architecture was generalized by integrating, in a single
model, the axial and appendicular movements of a centipede in the fastest gait pattern of locomotion.
1 INTRODUCTION
Central Pattern Generators (CPGs) are neural
circuits that can, without any sensory input, produce
rhythmic patterned outputs (Marder et alli, 1995).
These networks underlie the production, in a large
spectrum of species, of a wide variety of rhythmic
motor patterns such as walking, swimming or flying.
For that reason, the scientific community devotes
enormous efforts to full comprehend it and, as fast
as new biological explanations are proposed to
explain the mechanism underlying the functioning of
CPGs, several mathematically strict models are
developed with the purpose of encompass their
effects to fields like robotics, computing and
artificial intelligence.
The most common approach to the development
of models for CPGs is based on dynamical system
theory (Golubitsky et alli, 1997). Usually, the
behaviour of the neurons in CPGs is modelled
through the help of non-linear coupled oscillators.
As one may know, the strategies to solve those types
of systems cover a vast and sophisticated
mathematical ground governed by differential
equations. The difficulty to analyse those systems
increases even more when the biochemical processes
involved in the modelling of CPG activity are
considered. On the other hand, a discrete and
generalized model approach could produce the same
results with the advantage of modularity and quick
development without any lost of accuracy. In this
work, we intend to use one of these models to
reproduce the locomotion of a centipede, hoping to
demonstrate the power of such models.
A special class of topology-independent graph
dynamics called Scheduling by Multiple Edge
Reversal (SMER), developed initially with the
purpose of solve some problems in distributed
computing, present itself as an interesting way of
predict and reproduce the behavior of many
biological oscillatory neuronal networks.
58
R. Braga R., Yang Z. and M. G. França F. (2008).
IMPLEMENTING AN ARTIFICIAL CENTIPEDE CPG - Integrating Appendicular and Axial Movements of the Scolopendromorph Centipede.
In Proceedings of the First International Conference on Bio-inspired Systems and Signal Processing, pages 58-62
DOI: 10.5220/0001069800580062
Copyright
c
SciTePress
In the following sections we will try to briefly
explain the SMER algorithm and show how, starting
from it, we can develop a model for the inner
biological behavior of CPGs. After that, we will
hold some discussions on centipedes, its axial and
appendicular movements, and lastly, an
experimental model will be draw as much as the
conclusions.
2 SMER AND ARTIFICIAL CPGS
SMER is an algorithm used in Distributed and
Parallel Computation as a tool to allow a given
number of processes sharing a finite number of
resources among them, without the occurrence of
deadlock or starvation. SMER is a generalization of
the Scheduling Edge Reversal (SER) graph
dynamics. In order to understand SER, consider a
given number of processes and resources as part of a
neighbourhood-constrained system represented by
an acyclic graph. Processes are represented by nodes
and resources by oriented edges. Each node will be
in one of two possible states: operating or idle; also,
each edge will be always point to the process that
has the resource turn available to. So, when a node
has all the shared edges pointing towards it, i.e., has
all the resources turns available, it changes from the
idle state to the operating state (in this case, this
node is also called sink node). Once this operating
process has finished operation, it reverses all its
oriented edges to its neighbours. Although that is not
the purpose of this work, it’s possible to prove that if
the initial graph is acyclic, then no process will be
idle forever and, more importantly, the system will
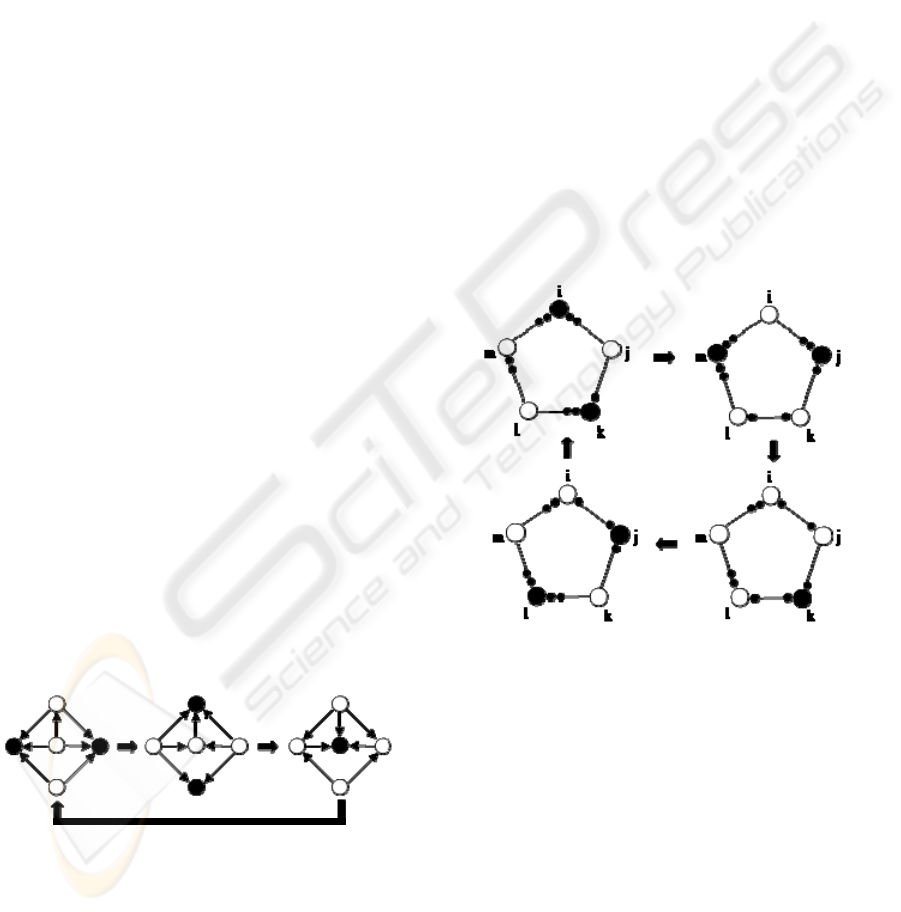
oscillate (see Figure 1). More than that, at any cycle
of oscillation, every process will operate the exactly
same number of times (Barbosa, 1996).
Figure 1: An example of the SER graph dynamics. Black
nodes represent operating processes; white nodes represent
idle processes.
Note that, even though the above described SER
mechanism is enough to solve much of the problems
of resource sharing, there is no differentiation
among the node’s time of task execution. It’s fair to
imagine that under certain circumstances some
processes will need of its shared resources for a
longer period of time than the others. To encompass
this scenario, the SMER algorithm was created as a
generalization of SER. In this new algorithm, all the
characteristics of the SER persist with the difference
that each node will have associated with it an natural
number r, called reversibility, and between any two
nodes is allowed to exist any number of oriented
edges. Once a node has pointing towards it, from all
of its neighbours, a number at least equal to its
reversibility, this node is allowed to operate. When
operation has finished, a node will reverse a number
of edges equal to his reversibility to all of its
neighbours (see Figure 2). Among the characteristics
of SMER, one very important is that for any system
with arbitrary reversibilities of its nodes, there is
always at least one possible periodic SMER
solution.
Figure 2: An example of a SMER graph. Note that to
avoid the existence of several arrows connecting two
nodes, a different representation of resource dependency is
adopted. In this example the reversibilities are i=l=m=2,
j=k=1.
Once we have defined what SMER is and how it
works, it’s important to clarify exactly how it
connects with CPGs. As said before, CPGs are the
underlying mechanism of a series of rhythmic
patterns of locomotion. Although it is not
completely clear how it exactly works, some
biological mechanisms have been found and are
credited as small units in the construction of CGPs.
One of those real neuronal mechanisms is called
post-inhibitory rebound (PIR) and is capable of
IMPLEMENTING AN ARTIFICIAL CENTIPEDE CPG - Integrating Appendicular and Axial Movements of the
Scolopendromorph Centipede
59

produce an alternate cycle of activity in a group of
inhibitory neurons in the absence of external
stimulus (Pirtle and Satterlie, 2007). Although the
PIR phenomenon is a complex subject, it is
interesting to note that it matches perfectly to the
mutual exclusion activity between neighbouring
nodes coupled under SMER. It will be the theory
behind the construction of modules that, in our
model, will act just like a set of interconnected
inhibitory neurons exhibiting PIR. These modules
will be called Oscillatory Building Blocks (OBBs).
So, instead of modeling electrophysiological
activities of interconnected neurons based on
membrane potential functions, we build an artificial
CPG network with SMER-based OBBs for the
exploration of the collective behaviour networks of
purely inhibitory neurons.
3 THE ARTIFICIAL CENTIPEDE
Centipedes form a very special species of
arthropods. They are capable of, combining axial
and appendicular movements, attaining great speed
with energetic efficiency. These unique
characteristics of the centipedes stimulate a great
number of biologists to study his static anatomy and
the kinematics of his locomotion leading to a great
amount of interesting information about this animal.
For instance, biologists thru the use of high-speed
cameras discovered that the number of legs touching
the ground at a high-speed movement decreases
when compared to the low-speed one , leading to a
bigger distance between the supporting legs. In the
extreme, a centipede can be supported for only four
legs. Also, there is a direct correlation between the
axial pattern of undulation and the speed.
Nevertheless, whatever the speed is, in each segment
contralateral legs will always step alternately
(Anderson, Shultz and Jayne, 1995). All this aspects
have to be taken into account while modeling the
centipede’s movement.
As a simple observation of a moving centipede
may suggest, the challenge is the integration
between two different components: the appendicular
and the axial. It’s reasonable to infer that a good way
of tackle this problem could be made through the
analysis of each movement separately, defining its
period and trying to construct a SMER-based OBB
for a later synchronization between the two.
Although it seems a good strategy, it lacks an
important aspect of the problem: the two types of
movements are connected in a much deeper level.
For example, it’s impossible to see a real centipede
to put two contralateral legs in any position different
that the one caused by alternately stepping.
Therefore, this approach would not reproduce that
subtle aspect of the locomotion of the centipede.
To correctly model the locomotion of a
centipede, with the maximum similarity to its
complex behavior, one has to construct the OBB
with eight nodes, i.e., motor neurons, enclosing one
whole segment. In this case, the network responsible
for the connection of these OBBs has to be one that
follows the full length of the animal, from the
anterior to the posterior segment. But before we see
in detail the whole model, let’s see more of each
centipede’s movement as a way to understand how
this OBB will be made and how the connections
among them will be put. Consider in the following a
scolopendromorph centipede in the fastest pattern
gait of locomotion, i.e., the amplitude of lateral
bending has the largest value and the fastest speed of
dislocation is attained. Also, it is important to note
that this kind of centipede has 21 leg bearing
segments linked by flexible membranes serving as
the only intersegmental articulation.
3.1 Appendicular Movement
As said before, in any given speed of the centipede,
two legs from the same segment are always in
opposite positions, i.e., when the left leg of a
segment is flexing the other in that segment is
extending. Also, it is important to note that the legs
that are in the concave side of an undulating wave
are always extending. The last statement is the most
important one since ties the axial and the
appendicular movements.
For the sake of simplicity and without any loss
of generality, let’s assume the appendicular
movement being defined as the action of two
antagonic muscles: flexor and extensor. The first one
is responsible for lifting a leg from the ground and
the later one for doing the opposite. In this
simplification, let’s also assume that when a leg is
touching the ground it is also pushing it backwards,
allowing the effective movement of the animal.
3.2 Axial Movement
In the fastest speed a centipede can attaining
approximately 1.5 times his length per second
(
1
5.1
−
Ls
) with a correspondents
Hzf 45.3=
and
11
=
λ
(Anderson, Shultz and Jayne, 1995). As a
result, we infer that each concave section of the
undulating wave it is composed for approximately 5
body segments. Also, for the sake of simplicity and
BIOSIGNALS 2008 - International Conference on Bio-inspired Systems and Signal Processing
60
without any lose of generality, let’s suppose the
lateral bending of a centipede as the result of a pair
of antagonic muscles: one causing the left and the
other causing the right bending.
4 EXPERIMENTAL
EXPLORATIONS
Our artificial centipede was designed to reproduce
the macroscopic features of its real counterpart.
Following the before mentioned characteristics and
simplifications, each segment will have six artificial
muscles: two pairs of extensors and flexors (one pair
per leg), one muscle responsible for the lateral
bending to the right and another for the left. As a
didactic resource, Figure 3 shows the schematic
representation of our artificial centipede’s segment
taking into the account the artificial muscles
mentioned before. The intersegmental articulation is
represented by a single pivot. Once more, note that
we consider that the extensor muscle is in action, the
respective leg is producing traction.
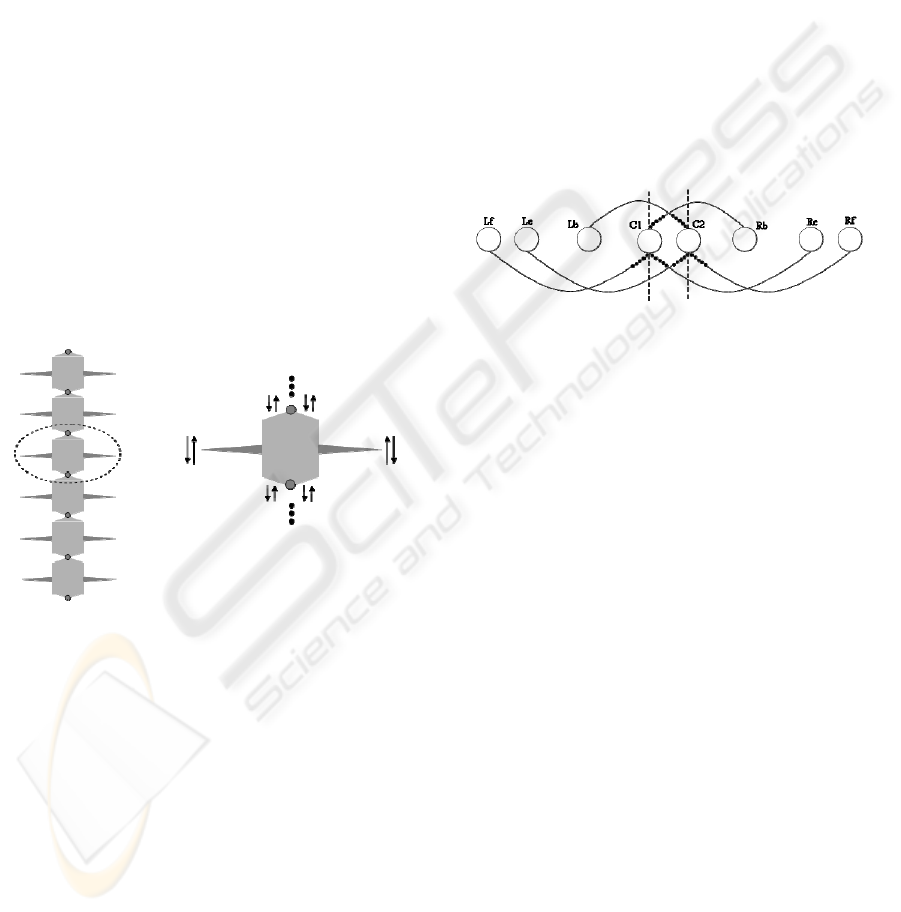
Figure 3: The Artificial Centipede design. In the left it is
displayed 6 of the 21 segments of the model. In the right,
the degrees of freedom in one segments is showed.
4.1 The Centipede OBB
As we saw before, to integrate the two types of
movements, the OBB has to enclose the whole
animal’s segment. So, in this model we have to
generate a SMER-based network capable of
reproducing all the intermediate positions that each
muscle assumed during the periodic movement. At
this point, all the information retrieved during the
analyses of real centipedes comes together.
In this OBB there are also two additional nodes,
represented in the middle of Figure 4, that are
responsible for the connection among the OBBs,
represented by the dotted line, and for the activation
of the others nodes, the artificial muscles. Note also
that the reversibility of those two connection-nodes
is 5, meaning that both of them are only activated
when each connected edge is fully directed to them.
Under another point of view, this also means that the
others nodes, the artificial muscles, will be activated
for a period of time five times longer them those
two, since its reversibility is one.
The above mentioned reversibility, i.e., r = 5,
was obtained from the analysis of the undulating
wave that covers the centipede from the anterior to
the posterior segment (see Figure 5.b). It is half of
the wavelength.
Figure 4: The resulting Oscillatory Building Block (OBB).
C1 and C2 are the inter-segment connection nodes; dotted
lines display the connection to other OBBs. Lb and Rb are
nodes representing the artificial motor neurons/muscles
responsible for the left and right bending, respectively. Le
and Re are nodes representing the artificial motor
neurons/muscles responsible for the extension of the left
and right legs, respectively. Finally, Lf and Rf are the
nodes representing the artificial muscles responsible for
the flexion of left and right legs, respectively.
4.2 The SMER Network
Now that the OBB is built, it is necessary to connect
them in a network that will reproduce the body
behaviour of the animal. Since the locomotion
pattern of the centipede is an undulating wave
covering the whole body, the design of the network
started with this perception and tried to reproduce
this characteristic. Fortunately, this proposition
proved correct and the SMER-based network,
responsible for the connection of the OBBs is one
that produces the activation of each OBB in the
same direction as the travelling wave (see Figure 5).
.
IMPLEMENTING AN ARTIFICIAL CENTIPEDE CPG - Integrating Appendicular and Axial Movements of the
Scolopendromorph Centipede
61
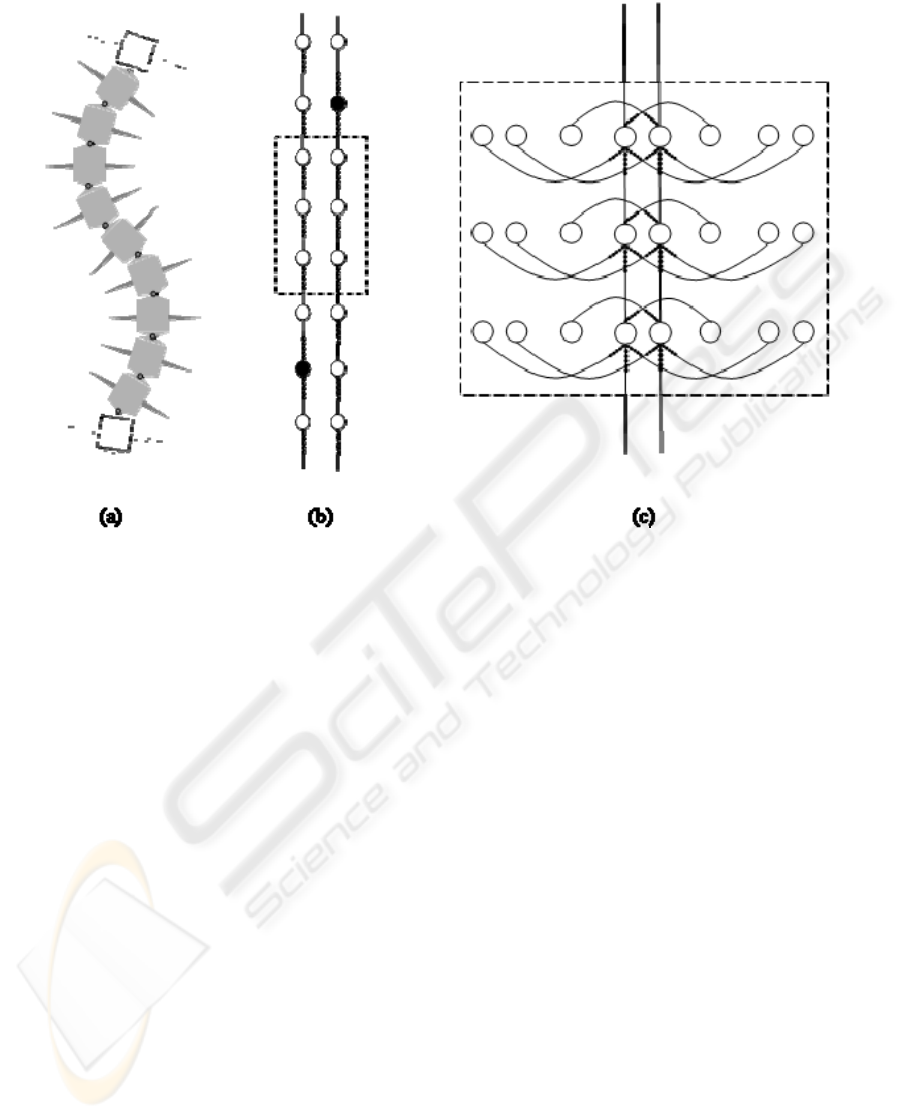
Figure 5: (a) The artificial centipede scheme (only 11 segments shown); (b) The functioning axial SMER-based network
(without the OBB details); (c) SMER-based OBBs (3 OBBs shown).
5 CONCLUSIONS
Since the beginning of the study of Central Pattern
Generators, one of the most critical problems was to
understand and to model the biological macroscopic
cyclic behaviour observed in terms of small
nonlinear units. As an alternative to the usual
continuous numerical methods applied in this field,
the use of a discrete and generalized model to mimic
the cyclic behaviour of CPGs was proposed in this
work. In this aspect, the use of distributed
algorithms avoids the usual complexity of the usual
approach without losing expressivity or generality.
The present work shows the application of one
of these algorithms (SMER) to model the complex
locomotion of a centipede at its fastest gait pattern
speed. Although others ways of reaching that
objective may exist, we believe that our approach
showed significant advantages in aspects like time
consumed, facility and acceptable correlation with
the reality. We believe that the strategy adopted in
this work could help biologists and
neurophysiologists to not only test the current
theories in Central Pattern Generator’s functioning,
but also develop new points of view in the
construction of complete explanations to the
phenomenon of the generation of rhythmic patterns
in animals.
REFERENCES
Barbosa, V.C., 1996. An Introduction to Distribuited
Algorithms, The MIT press, 1
nd
edition.
Marder, E., Nandim, F., Calabrese, R.L., 1995. Principles
of rhythmic Motor Pattern Production, Physiological
Reviews.
Anderson, B.D., Shultz, J.W., Jayne, B.C., 1995. Axial
Kinematics and muscle activity during terrestrial
locomotion of the centipede Scolopendra Heros. The
Company of Biologists Limited.
Golubitsky, M., Stewart, I., Buono, P., Collins, J.J., 1997.
A modular network for legged locomotion. N.H.
Elsevier.
Pirtle, T. J., Satterlie, R.A., 2007. The role of post-
inhibitory rebound in the locomotor central-pattern
generator of Clione limacine, Integr. Comp. Biol.
Yang, Z., França, F.M.G., 2003. A generalized locomotion
CPG architecture based on oscillatory building blocks,
Biol. Cybern., Springer-Verlag.
BIOSIGNALS 2008 - International Conference on Bio-inspired Systems and Signal Processing
62
