
ACCURACY ANALYSES OF PASSIVE TRACKING OF SEVERAL
CLICKING SPERM WHALES
A Case of Complex Sources Binding
Fr´ed´eric Caudal and Herv´e Glotin
System & Information Sciences Laboratory (LSIS - UMR CNRS 6168), Universit´e du Sud Toulon Var
BP 20132, 83957 La Garde Cedex, France
Keywords:
Delay estimation, marine mammals, acoustic tracking, Cram´er-Rao Lower Bound, acoustic propagation.
Abstract:
This paper provides a real-time passive underwater acoustic method to track multiple emitting whales using
four or more omni-directional widely-spaced bottom-mounted hydrophones and to evaluate the performance
of the system via the Cram´er-Rao Lower Bound (CRLB) and Monte Carlo simulations. After a non-parametric
Teager-Kaiser-Mallat signal filtering, rough Time Delays Of Arrival are calculated, selected and filtered, and
used to estimate the positions of whales for a constant or linear sound speed profile. The complete algorithm
is tested on real data from the NUWC and the AUTEC. The CRLB and Monte Carlo simulations are computed
and compared with the tracking results. Our model is validated by similar results from the US Navy and
Hawaii univ labs in the case of one whale, and by similar whales counting from the Columbia univ. ROSA
lab in the case of multiple whales. At this time, our tracking method is the only one giving typical speed and
depth estimations for multiple (5) emitting whales located at 1 to 5 km from the hydrophones.
1 INTRODUCTION
Processing of Marine Mammal (MM) signals for pas-
sive oceanic acoustic localization is a problem that
has recently attracted attention in scientific litera-
ture and in some institutes like the AUTEC
1
and
the NUWC
2
. Motivation for processing MM signals
stems from increasing interest in the behavior of en-
dangered MM
3
. One of the goals of current research
in this field is to develop tools to localize the vocal-
izing and clicking whale for species monitoring. In
this paper we propose a low cost time-domain track-
ing algorithm based on passive acoustics. The exper-
iments of this paper consist in tracking an unknown
number of sperm whales (Physeter catodon). Clicks
are recorded on two datasets of 20 and 25 minutes
on a open-ocean widely-spaced bottom-mounted hy-
drophone array. The output of the method is the
track(s) of the MM(s) in 3D space and time. In Sec-
tion 2 we briefly review studies of source separation
methods and the main characteristics of MM signals
and we propose a real-time algorithm for MM tran-
1
Atlantic Undersea Test & Evaluation Center - Bahamas
2
Naval Undersea Warfare Center of the US Navy
3
This work is founded by the ”Conseil r´egional
Provence-Alpes-Cˆote d’Azur” France, and partly by Chrisar
Software Inc.
sient call localization. In Section 3, we recall briefly
the CRLB (Kay, 1993) and the confidence ellipses
theory. In Section 4 we show and compare results of
tracks estimates with results from specialized teams
and the performance system with the CRLB.
2 PROBLEM FORMULATION
This papers deals with the 3D tracking of MM using
a widely-spaced bottom-mounted array in deep wa-
ter. It focuses on sperm whale clicks; detection and
classification are not a concern. There were previous
algorithms developed in the state of art (Giraudet and
Glotin, 2006b; Giraudet and Glotin, 2006a; Nosal and
Frazer, 2006; Morrissey et al., 2006) but none are able
to have satisfying results for multiple tracks. Most of
them are far from being real-time. The main goal is
to build a robust and real-time tracking model, despite
ocean noise, multiple echoes, imprecise sound speed
profiles, an unknown number of vocalizing MM, and
the non-linear time frequency structure of most MM
signals. Background ocean noise results from the ad-
dition of several noises: sea state, biological noises,
ship noise and molecular turbulence. Propagation
characteristics from an acoustic source to an array of
55
Caudal F. and Glotin H. (2008).
ACCURACY ANALYSES OF PASSIVE TRACKING OF SEVERAL CLICKING SPERM WHALES - A Case of Complex Sources Binding.
In Proceedings of the International Conference on Signal Processing and Multimedia Applications, pages 55-62
DOI: 10.5220/0001933700550062
Copyright
c
SciTePress
hydrophones include multipath effects (and reverber-
ations), which create secondary peaks in the Cross-
Correlation (CC) function that the generalized CC
methods cannot eliminate.
2.1 Material
Table 1: Hydrophones positions: D=Datasets,
Dist=Distance to barycenter (m).
D Hydros Dist X (m) Y (m) Z (m)
D1
H 1 5428 18501 9494 -1687
H 2 4620 10447 4244 -1677
H 3 2514 14119 3034 -1627
H 4 1536 16179 6294 -1672
H 5 3126 12557 7471 -1670
H 6 4423 17691 1975 -1633
D2
H 7 1518 10658 -14953 -1530
H 8 4314 12788 -11897 -1556
H 9 2632 14318 -16189 -1553
H 10 3619 8672 -18064 -1361
H 11 3186 12007 -19238 -1522
The signals are records from the ocean floor near An-
dros Island - Bahamas (Tab.1), provided with celer-
ity profiles and recorded in March 2002. Datasets
are sampled at 48 kHz and contain MM clicks and
whistles, background noises like distant engine boat
noises. Dataset1 (D1) is recorded on hydrophones
1 to 6 with 20 min length while dataset2 (D2)
is recorded on hydrophones 7 to 11 with 25 min
length. We will use a constant sound speed with
c = 1500ms
−1
or a linear profile with c(z) = c
0
+ gz
where z is the depth, c
0
= 1542ms
−1
is the sound
speed at the surface and g = 0.051s
−1
is the gradi-
ent. Sound source tracking is performed by continu-
ous localization in 3D using Time Delays Of Arrival
(T) estimation from four hydrophones.
2.2 Signal Filtering
A sperm whale click is a transient increase of sig-
nal energy lasting about 20 ms (Fig.1-a). Therefore,
we use the Teager-Kaiser (TK) energy operator on the
discrete data:
Ψ[x(n)] = x
2
(n) − x(n+ 1)x(n− 1), (1)
where n denotes the sample number. An important
property of TK is that it is nearly instantaneous given
that only three samples are required in the energy
computation at each time instant. Considering the raw
signal s(n) as:
s(n) = x(n) + u(n),
where x(n) is the signal of interest (clicks), u(n) is an
additive noise defined as a process realization consid-
ered wide sense stationary (WSS) Gaussian during a
short time, by applying TK to s(n), Ψ[s(n)] is:
Ψ[s(n)] ≈ Ψ[x(n)] + w(n),
where w(n) is a random gaussian process (Kandia
and Stylianou, 2006). The output is dominated by
the clicks energy. Then, we reduce the sampling
frequency to 480Hz by the mean of 100 adjacent
bins to reduce the variance of the noise and the data
size. We apply the Mallat’s algorithm (Mallat, 1989)
with the Daubechies wavelet (order 3). We chose
this wavelet for its great similarity to the shape of
a decimated click (Giraudet and Glotin, 2006b; Gi-
raudet and Glotin, 2006a). The signal is denoised
with a universal thresholding (Donoho, 1995) defined
as D(u
k
,λ) = sgn(u
k
)max(0,|u
k
| − λ), with u
k
the
wavelet coefficients, λ =
p
(2log
e
(Q))σ
N
σ
˜
N
, and Q
the length of the signal resolution level to denoise.
The noise standard deviation σ
N
is calculated on each
10s window on the raw signal with a maximum like-
lihood criterion. σ
˜
N
is the standard deviation of the
wavelet coefficients on a resolution level of a gener-
ated, reduced and 0-mean Gaussian noise. This fil-
tering step is very fast without any parameter. Fig.1-
d is the filtered signal on multiple (Fig.1-c) emitting
MMs.
2.3 Rough
e
T Estimation
First, T estimates are based on MM click realignment
only. Every 10s, and for each pair of hydrophones
(i, j), the difference between times t
i
and t
j
of the ar-
rival of a click train on hydrophones i and j is referred
as T(i, j) = t
j
−t
i
. Its estimate
e
T(i, j) is calculated by
CC of 10s chunks (2s shifting) of the filtered signal
for hydrophones i and j (Giraudet and Glotin, 2006b;
Giraudet and Glotin, 2006a). We keep the 35 high-
est peaks on each CC to determine the corresponding
e
T(i, j). The filtered signals give a very fast rough esti-
mate of T (precision ± 2ms). Fig.(1.e) shows the CC
with the raw signal and (1.f) with the filtered signal.
Without filtering, CC generates spurious delays esti-
mates and the tracks are not correct. The maximum
e
T rank (Fig.2) in D1, pitching the source localization,
are high among the 35
e
T kept in the CC which justi-
fies this number.
2.4
e
T Selection and Localization with a
Constant Profile
Each signal shows echoes for each click (Fig.1 b),
maybe due to the reflection of the click train off the
SIGMAP 2008 - International Conference on Signal Processing and Multimedia Applications
56
2.32 2.33 2.34 2.35 2.36 2.37 2.38 2.39
−0.4
−0.2
0
0.2
0.4
Time of recording (s)
a
2 4 6 8 10
−0.5
0
0.5
1
b
Time of recording (s)
4 6 8 10 12
−0.5
0
0.5
1
Time of recording (s)
c
−8 −6 −4 −2 0 2 4 6 8
−10
0
10
e
4 6 8 10 12
0
0.5
1
Time of recording (s)
d
−8 −6 −4 −2 0 2 4 6 8
0
5
10
f
CC: delay estimation
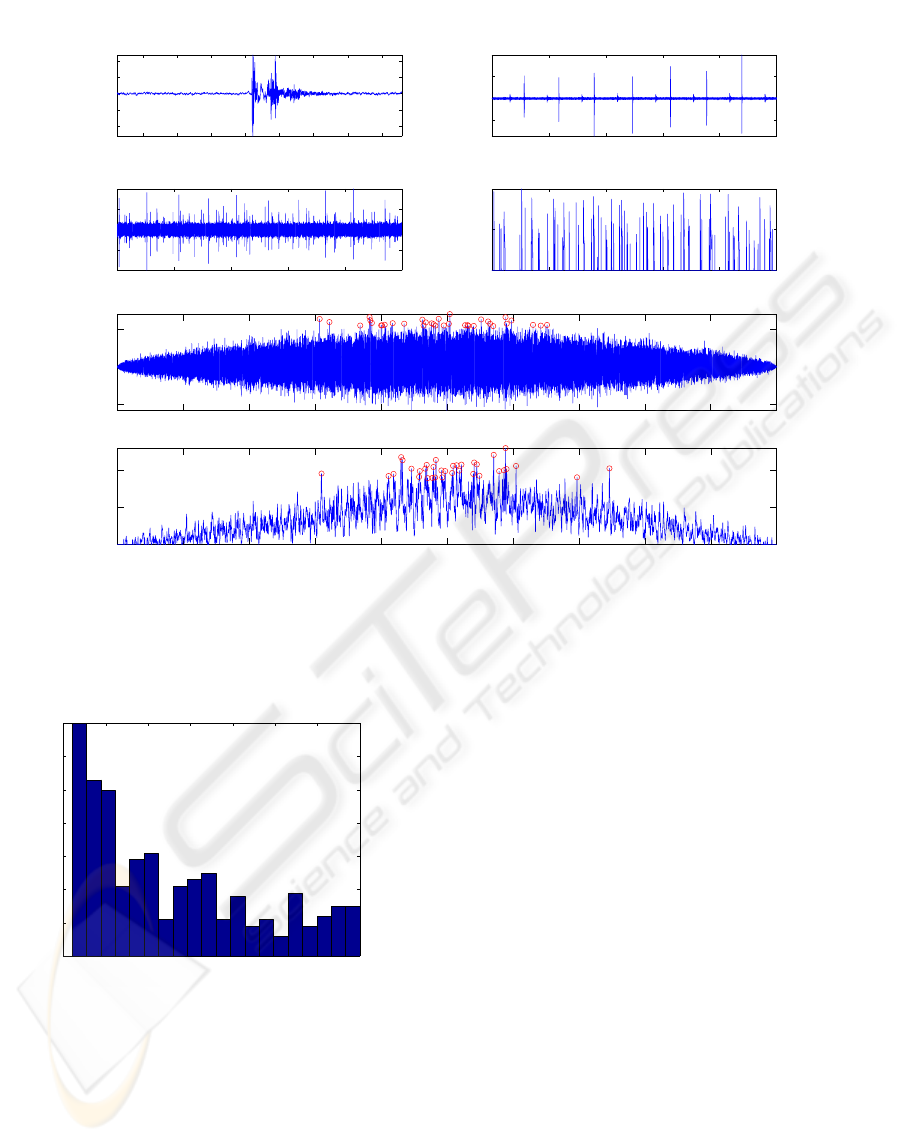
Figure 1: (a): Detail of a click. (b): raw signal (D2) of hydrophone 7 (H7) during the first 10s of recording, containing 7
clicks and their echoes. (c): raw signal (D1) of H3 during the first 10s of recording showing multiple emissions. (d): (c) after
filtering. (e): CC between (c) and corresponding raw signal chunk of H1. (f): idem than (e) but with (d). Circles correspond
to the 35 maximum peaks.
0 5 10 15 20 25 30 35
0
10
20
30
40
50
60
70
Number of
˜
T
Maximum
˜
T rank
Figure 2: Maximum
e
T rank histogram in the CC for each
triplet.
ocean surface or bottom or different water layers. We
use a method based on autocorrelation (Giraudet and
Glotin, 2006b; Giraudet and Glotin, 2006a) to elimi-
nate it. Then, thanks to the
e
T transitivity system de-
scribed in (Giraudet and Glotin, 2006b) we keep
e
T
triplets coming from the same source. Finally, thanks
to the measured delays and an acoustic model based
on a constant sound speed profile, the least squares
cost function determines the MM positions using a
multiple non linear regression with Gauss-Newton
method (Levenberg-Marquardt)(Giraudet and Glotin,
2006b). The residuals are approximatively following
a Chi-square distribution with Nc− d degrees of free-
dom, noted X
2
Nc−d
, Nc is the number of hydrophones
couples considered and d the number of unknowns,
here 3 (x,y,z). The position is accepted if the residual
is inferior to a threshold x, That is calculated solving
P = prob(X
2
Nc−d
> x) with P = 0.01 (we keep 99% of
the estimates).
2.5 Source Localization with a Linear
Sound Speed Profile
It is well known that the ray paths in a medium with
linear sound speed profile are arcs of circles and fur-
ther the radius of the circle can be computed (Fig.3).
c
s
is the sound speed at the source, θ
s
is the launch
angle of the ray at the source, measured relatively to
the horizontal. From the geometry shown in Fig.3, the
center of the circle, (xc,zc), along which the ray path
is an arc, is (White et al., 2006):
ACCURACY ANALYSES OF PASSIVE TRACKING OF SEVERAL CLICKING SPERM WHALES - A Case of
Complex Sources Binding
57
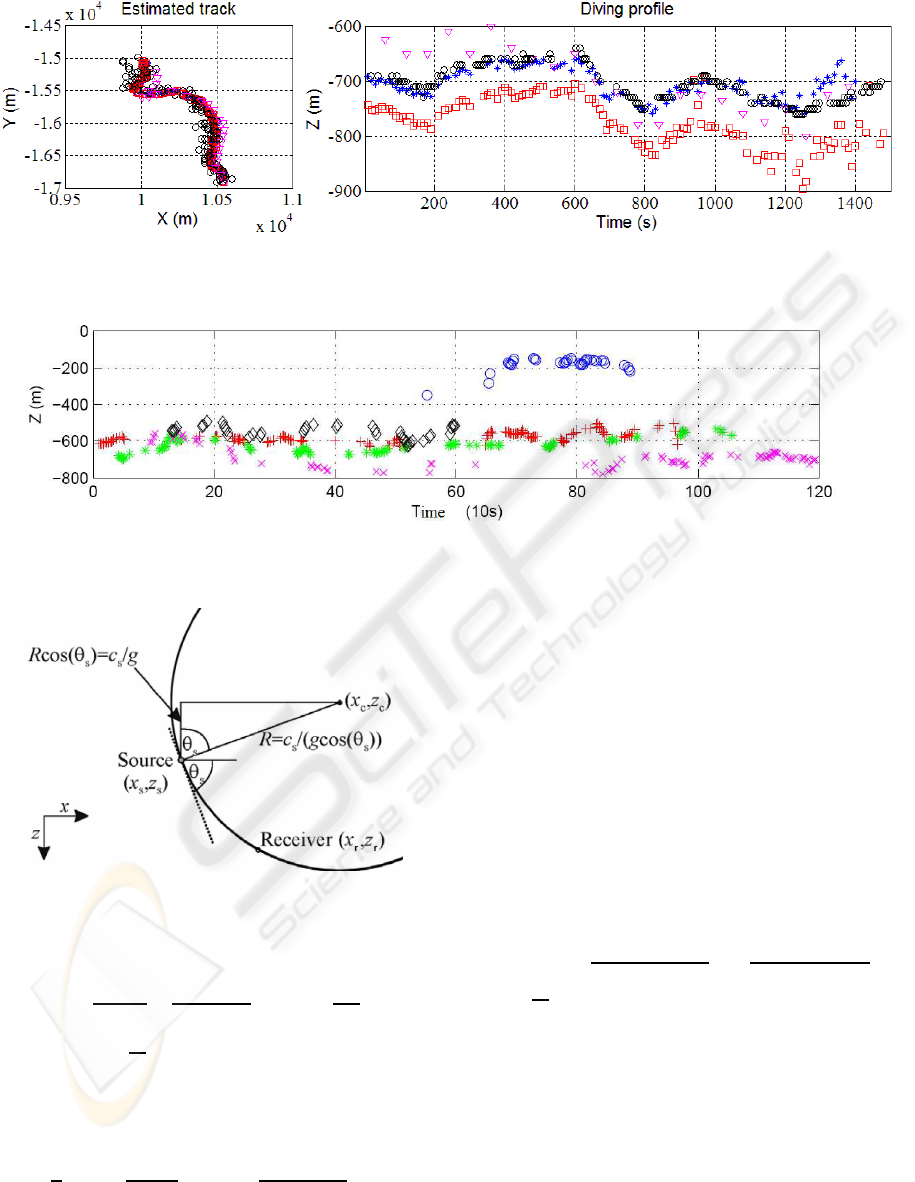
Figure 4: Plan view and diving profile of the MM in D2, our estimates with a linear (A) (Caudal and Glotin, 2008) or constant
profile (2); and estimates from Morrissey’s (Morrissey et al., 2006) (▽) and from Nosal’s (Nosal and Frazer, 2006) methods
(o).
Figure 5: Averaged diving profile in D1. Each symbol corresponds to one whale in Fig.6 (Bottom to top (+), (o), (A), (x),
(⋄)).
Figure 3: Geometry for a source and receiver in a linear
profile (White et al., 2006).
x
c
=
x
s
+ x
r
2
+
(z
s
− z
r
)
2(x
s
− x
r
)
(z
r
− z
s
+
2c
s
g
),
z
c
= z
s
−
c
s
g
.
(2)
For linear sound speed profile the course time τ of
the ray is:
τ =
1
g
log
z
c
− z
s
z
c
− z
r
− log
R+ x
c
− x
s
R+ x
c
− x
r
. (3)
Using Eqs.(2)-(3) allows one to compute the propaga-
tion time from the source to any receiver and then the
whale position.
3 THE CRLB AND CONFIDENCE
REGIONS
3.1 The CRLB
The CRLB provides the maximum acurracy for the
estimation of the position. Considering a constant
sound speed profile, the function model of the TDOA
is defined:
s(θ) =
1
c
s
v
u
u
t
3
∑
k=1
(X
i,k
− θ
k
)
2
−
v
u
u
t
3
∑
k=1
(X
j,k
− θ
k
)
2
.
where X
{
i, j}
is the vector coordinate of hy-
drophone {i, j}, θ is the unknown parameters vec-
tor [x y z]
T
and c
s
the celerity. Thus, considering the
TDOA noise Gaussian and B its variance-covariance
matrix, the Fisher Information matrix is:
I
θ
= ∇
θ
s(θ)B
−1
∇
T
θ
s(θ).
Then, the CRLB is B
θ
= I
−1
θ
.
SIGMAP 2008 - International Conference on Signal Processing and Multimedia Applications
58
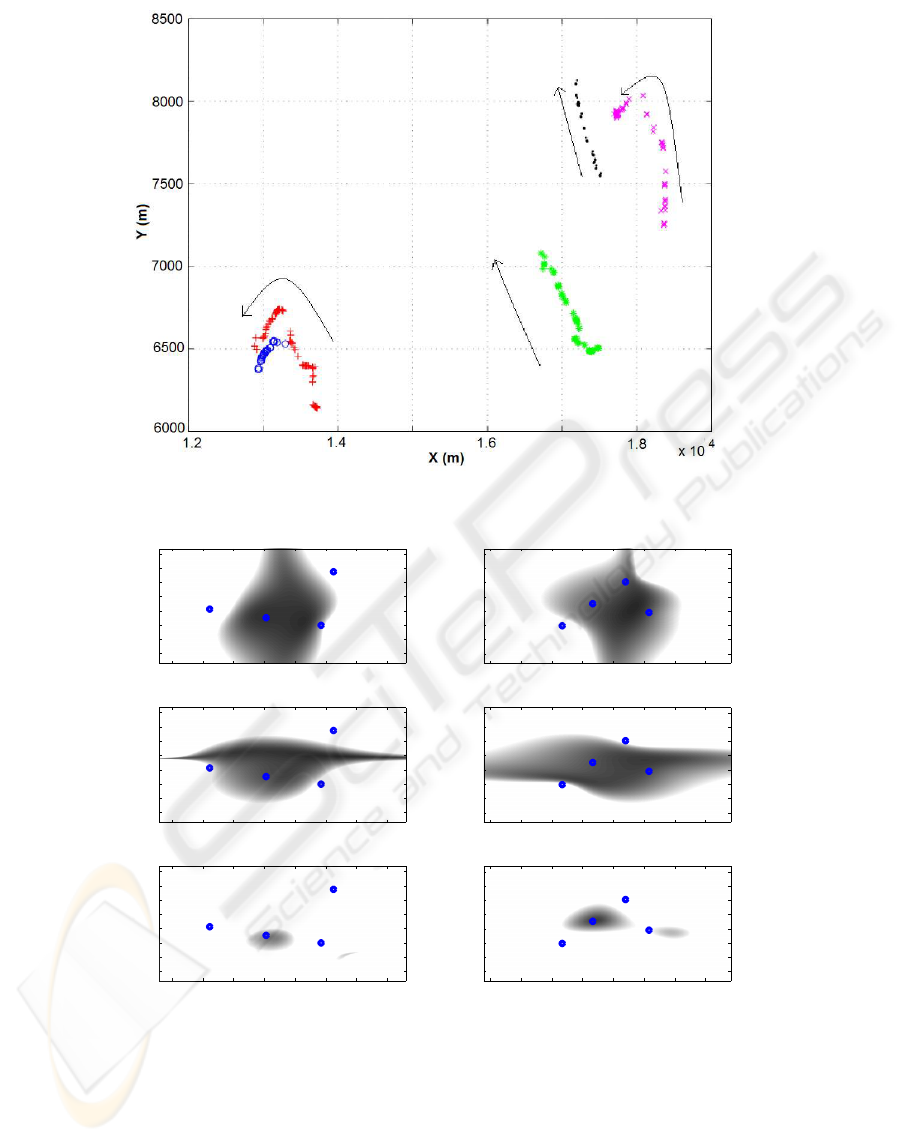
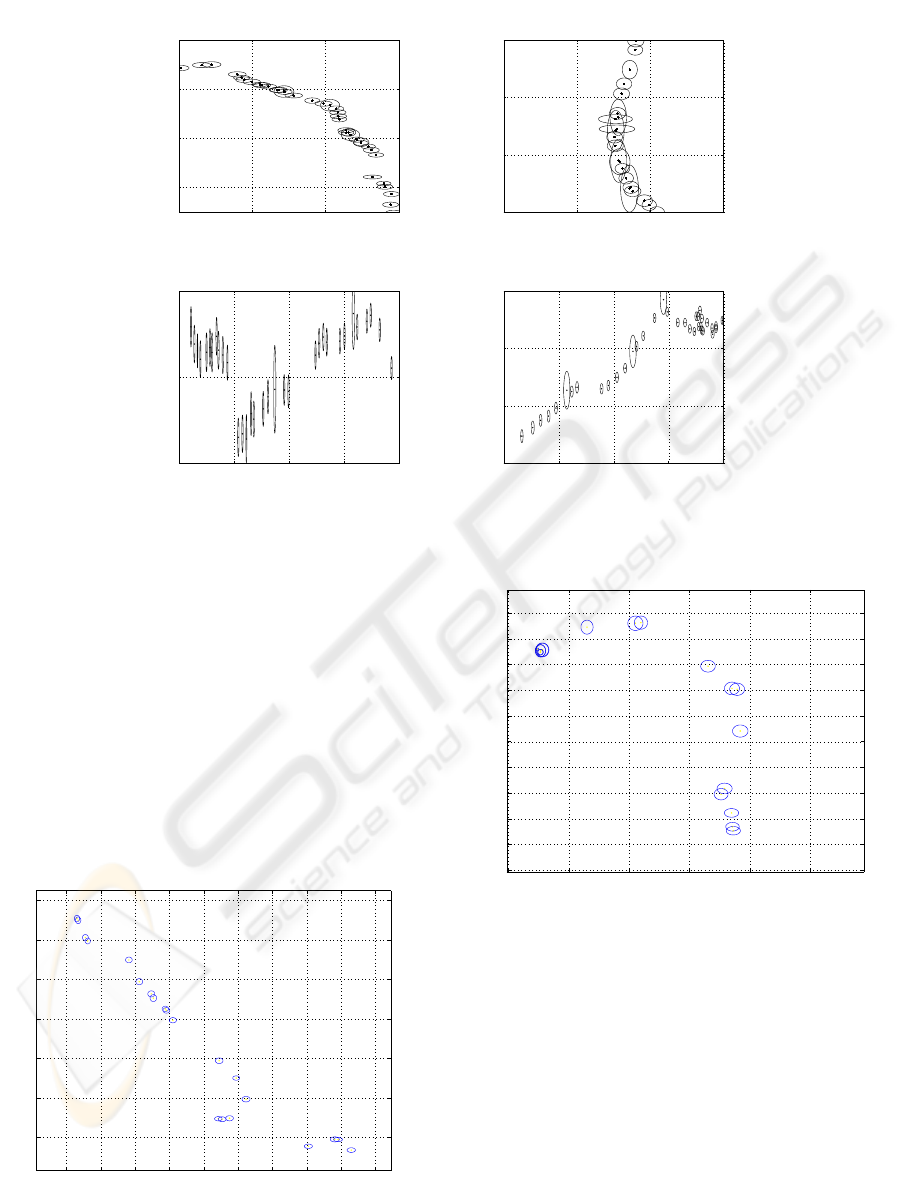
Figure 6: Plan view in D1. Each symbol corresponds to one of the 5 whales. The arrows stress the directions.
H1
H2
H3
H6
A
X (m)
Y (m)
0.8 1 1.2 1.4 1.6 1.8 2 2.2
x 10
4
−2000
0
2000
4000
6000
8000
10000
12000
H1
H2
H3
H6
B
X (m)
Y (m)
0.8 1 1.2 1.4 1.6 1.8 2 2.2
x 10
4
−2000
0
2000
4000
6000
8000
10000
12000
H1
H2
H3
H6
C
X (m)
Y (m)
0.8 1 1.2 1.4 1.6 1.8 2 2.2
x 10
4
−2000
0
2000
4000
6000
8000
10000
12000
H7
H8
H9
H10
D
X (m)
Y (m)
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8
x 10
4
−2.2
−2
−1.8
−1.6
−1.4
−1.2
−1
−0.8
x 10
4
H7
H8
H9
H10
E
X (m)
Y (m)
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8
x 10
4
−2.2
−2
−1.8
−1.6
−1.4
−1.2
−1
−0.8
x 10
4
H7
H8
H9
H10
F
X (m)
Y (m)
0.4 0.6 0.8 1 1.2 1.4 1.6 1.8
x 10
4
−2.2
−2
−1.8
−1.6
−1.4
−1.2
−1
−0.8
x 10
4
Figure 7: CRLB values scaled in gray colors with a plan view: black means a null CRLB and white a CRLB≥10 m. For all
the figures, a depth of 500m was chosen. (A): CRLB on x values, dataset D1. (B): y, D1. (C): z, D1. (D): x, D2. (E): y, D2.
(F): z, D2.
3.2 Confidence Regions
The confidence ellipse generalizes the notion of con-
fidence interval in case of gaussian vector in two di-
mensions. Considering a stochastic gaussian vector
X ∼ N(µ,Σ) of two dimensions with Σ > 0. We have
to determine the level curves of the gaussian distribu-
tionl, i.e. the points x of the ellipse
(x− µ)
∗
Σ
−1
(x− µ) = r
ACCURACY ANALYSES OF PASSIVE TRACKING OF SEVERAL CLICKING SPERM WHALES - A Case of
Complex Sources Binding
59

Being a symmetrical positive matrix, Σ can be
spectraly factorized as Σ = PDP
∗
where P is the or-
thonormal matrix of the eigen vectors and D the diag-
onal matrix of the eigen values. For each x, the point ˜x
is defined with ˜x = D
−1/2
Px, then | ˜x|
2
= r, i.e. all the
˜x is the circle centered with radius r : ˜x(ρ) = r[cos(ρ)
sin(ρ)]
∗
for 0 ≤ ρ < 2π. The parametric equation of
the ellipse is:
x(ρ) = µ+ rP
∗
D
1/2
cos(ρ)
sin(ρ)
,0 ≤ ρ < 2π
The quadratic norm of the stochastic vector
˜
X =
D
−1/2
PX is distributed with the chi-square distribu-
tion with 2 degrees of freedom. Putting r = 5.99, then
P(X ∈ x ∈ R
2
: (x− µ)
∗
Σ
−1
(x− µ) ≤ r) = 0.95
This means the real position is 95% likely to be inside
the ellipse.
4 RESULTS
4.1 Tracking Comparisons
For D2 (Fig.4), a constant and a linear sound speed
profile were used (Caudal and Glotin, 2008) and the
results are similar with the Morrissey’s (Morrissey
et al., 2006) and Nosal’s (Nosal and Frazer, 2006)
methods. The diving profile underlines a bias of about
50 to 100m between the linear and the constant pro-
files results, emphasizing the importance of the cho-
sen profiles. Moreover with the linear sound speed,
the results are about the same as Morrissey’s and
Nosal’s, who used profiles corresponding to the pe-
riod and place of the recordings. Results on D1 are
shown in Fig.6-5 for a linear sound speed profile. We
thus localize 5 MM. The confidence regions are com-
puted for the two datasets with a Monte Carlo method.
The ellipses maxima (30m) fit with MM length (20m).
4.2 CRLB Computation
The CRLB provide the maximum acurracy for the lo-
calization. Here, we computed the CRLB (in me-
ter) in the space (x,y,z) and plot the values for both
datasets (Fig.7). We consider that the standard devi-
ation of the noise is equal to the quantification noise
with a sampling frequency of 480Hz. The main de-
pendencies of the bounds are the noise and the array
configuration.
In figures 8 and 9, the CRLB on y and z is shown
for a depth of 1000m, and is just about the same for
H1
H2
H3
H6
B
X (m)
Y (m)
0.8 1 1.2 1.4 1.6 1.8 2 2.2
x 10
4
−2000
0
2000
4000
6000
8000
10000
12000
Figure 8: CRLB on y axis, plan view, dataset D1,
depth=1000m.
H1
H2
H3
H6
C
X (m)
Y (m)
0.8 1 1.2 1.4 1.6 1.8 2 2.2
x 10
4
−2000
0
2000
4000
6000
8000
10000
12000
Figure 9: CRLB on z axis, plan view, dataset D1,
depth=1000m.
a depth of 500m. Thus, as the mean depth of a sperm
whale is in this range, the CRLB likewise.
4.3 The Confidence Regions
We apply a Monte Carlo method with a gaussian dis-
tribution noise with the standard deviation described
above. For each
e
T realization, the source position
is calculated. We deduce the variance and the mean
for each position to plot the confidence regions with
a confidence level of 0.95, which means that there
is 0.95 probability for the whale to be in the ellipse
centered on the position. In D2, the mean values of
the confidence intervals on X, Y, Z axes are about
18, 16 and 30 m (Fig.10). This justifies the decima-
tion on the raw signal, because the error on X and
Y axes are close to the sperm whale length (20m).
The results confirm that the errors on the vertical axis
SIGMAP 2008 - International Conference on Signal Processing and Multimedia Applications
60
1.02 1.03 1.04 1.05
x 10
4
−1.6
−1.58
−1.56
−1.54
x 10
4
X (m)
Y (m)
1.03 1.04 1.05 1.06
x 10
4
−1.67
−1.66
−1.65
−1.64
x 10
4
X (m)
Y (m)
−1.68−1.66−1.64−1.62 −1.6
x 10
4
−800
−750
−700
Y (m)
Z (m)
−1.59−1.58−1.57−1.56−1.55
x 10
4
−800
−750
−700
−650
Y (m)
Z (m)
Figure 10: Confidence regions projection on X and Y and on Z and Y axes for D2 trajectory.
are meaningfully higher than the other axes because
the distance between each hydrophones in this direc-
tion (maximum difference on the Z axis between hy-
drophones is 200m) is smaller. The D1 results ob-
tained with a linear profile (Fig.6), indicate five tra-
jectories. The farthest whales in D1 from the hy-
drophones array center have a larger uncertainty with
an error of about 20 to 30m on X and Y axes (Fig.12),
while the whales close to the center (Fig.11) exhibit
an error of about 10 to 20m like for D2 (Fig.6). Those
uncertainties are reasonable according to the sperm
whale length.
1.67 1.68 1.69 1.7 1.71 1.72 1.73 1.74 1.75 1.76
x 10
4
6500
6600
6700
6800
6900
7000
7100
X (m)
Y (m)
Figure 11: Confidence regions projection on X and Y for
the whale in the middle (* symbol), D1 trajectory.
1.76 1.78 1.8 1.82 1.84 1.86
x 10
4
7000
7100
7200
7300
7400
7500
7600
7700
7800
7900
8000
X (m)
Y (m)
Figure 12: Confidence regions projection on X and Y for
the whale in the right (x symbol), D1 trajectory.
4.4 Performance Comparisons
Comparing the CRLB and the ellipses, we denote the
correlation between maximum accuracy and confi-
dence regions. Figures 7-C and 7-F show the accuracy
on z axis is ≥10 m for both datasets in the tracking
regions which is consistent with the ellipses results.
The CRLB (in D1, figures 6-D,E) also explains that
the farthest whale has larger confidence regions. But
for both datasets, the CRLB on x and y is far inferior
to 10m inside the array whereas our ellipses are about
10 to 20m. Maybe other parameters like the approxi-
ACCURACY ANALYSES OF PASSIVE TRACKING OF SEVERAL CLICKING SPERM WHALES - A Case of
Complex Sources Binding
61

mated celerity profile are involved in.
5 CONCLUSIONS
The tracking algorithm presented in this paper is real-
time on a standard laptop and works for one or mul-
tiple emitting sperm whales. Depth results with con-
stant speed contains a bias errors due to the refraction
of the sound paths from the MM to the receivers what
a linear speed corrects. An other way to tackle the
speed profile issue (Glotin et al., 2008) is to estimate it
as a fourth unknown in the regression. Our algorithm
has no species dependency as long as it processes
all transients. At this time, only our algorithm gives
results with typical speed and depth estimations for
multiple emitting whales. In D2, results indicate that
only one sperm whale was present in the area, unless
other whales in the area were quiet during the selected
25-min period. Moreover, according to ROSA Lab es-
timation based on click clustering, averaged number
of MM for each 5min chunks on D1 is [4.3;5.3;4;3.6]
(Halkias and Ellis, 2006) similar to ours ([4;4;4;3]).
The localization accuracy is computed via the CRLB
and the confidence ellipses which are correct consid-
ering the MM length. The high depth error is mainly
due to the low precision reachable with the CRLB
considering the array configuration. Our method pro-
vides thus robust online detecting/counting system of
clicking MM in open ocean.
REFERENCES
Caudal, F. and Glotin, H. (2008). Multiple real-time 3d
tracking of simultaneous clicking whales using hy-
drophone array and linear sound speed profile. In
ICASSP IEEE, volume 4p.
Donoho, D. L. (1995). De-noising by soft thresholding.
IEEE Trans. IT, 41:613–627.
Giraudet, P. and Glotin, H. (2006a). Echo-robust and real-
time 3d tracking of marine mammals using their tran-
sient calls recorded by hydrophones array. In ICASSP
IEEE.
Giraudet, P. and Glotin, H. (2006b). Real-time 3d tracking
of whales by echo-robust precise tdoa estimates with a
widely-spaced hydrophone array. Applied Acoustics,
67:1106–1117.
Glotin, H., Caudal, F., and Giraudet, P. (2008). Whales
cocktail party: a real-time tracking of multiple whales.
International Journal Canadian Acoustics, page 7p.
Halkias, X. and Ellis, D. (2006). Estimating the number
of marine mammals using recordings form one micro-
phone. In ICASSP IEEE.
Kandia, V. and Stylianou, Y. (2006). Detection of sperm
whale clicks based on the teager-kaiser energy opera-
tor. Applied Acoustics, 67:1144–1163.
Kay, S. (1993). Fundamentals of statistical signal process-
ing. PTR.
Mallat, S. (1989). A theory for multiresolution signal de-
composition: The wavelet representation. volume 11,
pages 674–693. IEEE Transaction on Pattern Analysis
and Machine Intelligense.
Morrissey, R., Ward, J., DiMarzio, N., Jarvisa, S., , and
Moretti, D. (2006). Passive acoustics detection and lo-
calization of sperm whales in the tongue of the ocean.
Applied Acoustics, 62:1091–1105.
Nosal, E. and Frazer, L. (2006). Delays between direct and
reflected arrivals used to track a single sperm whale.
Applied Acoustics, 62:1187–1201.
White, P., Leighton, T., Finfer, D., Powles, C., and Bau-
mann, O. (2006). Localisation of sperm whales
using bottom-mounted sensors. Applied Acoustics,
62:1074–1090.
SIGMAP 2008 - International Conference on Signal Processing and Multimedia Applications
62
