
USING PHYLLOTAXIS FOR DATE PALM TREE
3D RECONSTRUCTION FROM A SINGLE IMAGE
Ran Dror, Ilan Shimshoni
University of Haifa, Haifa, Israel
Keywords:
Phyllotaxis, 3D reconstruction, Palm tree.
Abstract:
Phyllotaxis is the study of the morphological order of plants. Remarkably, in spite of the overwhelming
diversity of plant morphology, there are common patterns that link a wide variety of species. The date palm,
having a phyllotactic order, possesses a simple, repetitive model. Only a small number of parameters are
needed to represent the phyllotactic order of the date palm. This a priori knowledge we have on the date
palm can help in the 3D reconstruction of the tree and can even make it possible to reconstruct a 3D model
from only one image. The proposed algorithm receives as input a single image of the date palm. Upon image
acquisition, the algorithm proceeds to search for, and locate, the trunk followed by a few prominent leaves.
From the location of the prominent leaves the algorithm proceeds to calculate tree model parameters, which
can then be used to search for additional, neighboring, leaves. Complete 3D reconstruction is achieved by
utilizing the calculated tree model parameters and by the known location of the leaves on the 2D image.
1 INTRODUCTION
Phyllotaxis
1
- a central area of research in plant mor-
phogenesis (Steeves and Sussex, 1989; Jean, 1994),
is the study of the arrangement of repeating units in
plants. For example, leaves around a stem, scales on
a pine cone or on a pineapple, the seeds in a sun-
flower head, etc. (see Figure 1). Remarkably, in spite
of the overwhelming diversity of plant morphology,
there are common patterns that link a wide variety of
species. Though the study of phyllotaxis is traced to
the first primitive observations in ancient times, it is
still a very active field today (Adler et al., 1997; Smith
et al., 2006; Reinhardt, 2005).
Within the three main types of phyllotactic pat-
terns found in nature, the most prevalent is the spiral
phyllotactic pattern. This pattern appears in the ma-
jority of the 250,000 or more species of higher plants
(Cummings and Strickland, 1998). The phyllotactic
patterns are formed at the microscopic level at the
growing tip of the plant - the apical meristem (see
Figure 2). Botanical units such as leaves and petals
are generated at the meristem as bulges of fast grow-
ing cells known as primordia. As more primordia de-
velop, they are pushed farther and farther from the
1
from Greek - phyllo (leaf) and taxis (order)
apex thus developing into the familiar features of the
plant, be it a leaf, a flower, or parts of a fruit. In spiral
phyllotaxis the angle between consecutive born pri-
mordia, called the divergence angle, is constant and
close to the Fibonacci angle of 137.5
◦
.
(a) (b) (c)
Figure 1: Examples of phyllotactic patterns in plants: (a) -
Echevaria subsseilis, (b) - Pineapple, (c) - Marguerite. (im-
ages from (Atela and Gol
´
e, 2008))
Spiral phyllotaxis is characterized by conspicuous
spirals, or contact parastichies, formed by sequences
of adjacent organs composing the structure (see Fig-
ure 2). Curiously, the numbers of parastichies run-
ning in opposite directions are usually two consecu-
tive Fibonacci numbers. The difference between age
indexes of two neighboring organs in the parastichy
288
Dror R. and Shimshoni I.
USING PHYLLOTAXIS FOR DATE PALM TREE 3D RECONSTRUCTION FROM A SINGLE IMAGE.
DOI: 10.5220/0001773802880296
In Proceedings of the Fourth International Conference on Computer Vision Theory and Applications (VISIGRAPP 2009), page
ISBN: 978-989-8111-69-2
Copyright
c
2009 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved

spiral indicates the number of parastichy spirals.
Figure 2: The apical meristem of A Norway spruce (Picea
abies) (Electron micrograph from (Rutishauser, 1988)). Pri-
mordia are numbered according to their age - the higher the
number, the older the primordium. Three of the eight right
parastichies spirals are illustrated in green, two of the thir-
teen left parastichies spirals are illustrated in purple. The
divergence angle between primordia six and seven which is
equal to the Fibonacci angle is marked in the center.
While phyllotactic order is common in the ar-
rangement of seeds in flowers and leaves around a
stem, the branching of the trunk and branches is al-
most unpredictable. In contrast to other trees, both
date and oil palms do not have branched trunks,
rather incorporating a single, non-branching, trunk
with leaves arranged in spiral phyllotactic order its
entire length.
This simple and consistent arrangement makes it
possible to describe these trees by means of a simple
model. The proposed algorithm uses the aforemen-
tioned model to reconstruct a 3D image of the tree.
It receives as input a single image of the date palm.
Upon image acquisition, the algorithm proceeds to
search for, and locate, the trunk followed by a few
prominent leaves. From the location of the promi-
nent leaves the algorithm calculates tree model pa-
rameters, which are then used to search for additional,
neighboring, leaves. The 3D reconstruction is done
using the tree model parameters and the location of
the leaves on the 2D image.
The palm tree was chosen because of its economic
importance and possible future practical implementa-
tions. There are two species of the palm family which
have economic importance, the oil palm and the date
palm. Palm oil is now the most widely produced veg-
etable oil in the world, demand for which is expected
to climb even further as one of the raw materials for
biodiesel. The date palm is one of the most impor-
tant economic tree crops that thrive in the desert re-
gions of the world. In order to reduce labour costs
and the risk of working at the top of the palm trees, a
number of robotic prototypes for maintenance of palm
trees were developed (Aracil et al., 1999; Ripin et al.,
2000; Shamsi, 1998). This study proposes a further
step in that it will include automatic vision algorithms
to guide these robotic systems.
2 RELATED WORK
The challenging problem of 3D reconstruction of
trees has been studied in (Shlyakhter et al., 2001;
Martinez et al., 2004). A number of calibrated im-
ages from different views of the tree are segmented
into the tree, and the background. The background
segments are discarded and the tree segments are then
compiled to reconstruct a 3D model resembling the
specific tree. The target of these works was render-
ing trees in virtual environments, whereas in our work
we try to measure the tree skeleton from only one
non calibrated image. Segmentation in these works is
done either manually (Shlyakhter et al., 2001), or for
cases where the tree background is distinct, such as
sky background (Martinez et al., 2004). More sophis-
ticated tree segmentation is done using 51 values mea-
sured at each pixel in (Haering and da Vitoria Lobo,
1999).
In a recent study (Teng et al., 2006) trunk structure
is extracted. The main challenge in this work is to deal
with the general trunk structure. In our case however,
the trunk is cylindrical.
Phyllotactic models are used in the field of com-
puter graphics for simulating realistic plant images.
An overview of these models is presented in Section
3. In a recent study (Kaewapichai et al., 2007) the
phyllotactic model was used to fit the arrangement of
the scales on a pineapple.
3 PALM TREE MODEL
Several mathematical phyllotactic models were devel-
oped by computer graphics researchers for the pur-
pose of simulating realistic plant images. Synthe-
sized images of plant structures with predominantly
flat, elongated or spherical geometry were created in
(Prusinkiewicz and Lindenmayer, 1990; Fowler et al.,
1989; Lintermann and Deussen, 1999) using three
USING PHYLLOTAXIS FOR DATE PALM TREE 3D RECONSTRUCTION FROM A SINGLE IMAGE
289

models: the planar model, the cylindrical model, and
the spherical model. These models relate phyllotaxis
to the packing problem of equally-sized organs.
Many phyllotactic plants do not fit into these mod-
els. Phyllotactic plants with a variety of organ sizes
and surface shapes were dealt with in (Fowler et al.,
1992). In this model plant organs are procedurally
placed on the plant’s surface.
The palm tree has a spiral phyllotactic pattern.
Characteristics of the date palm phyllotaxis have been
studied in (Elhoumaizia et al., 2002; Ferry, 1998). In
(Elhoumaizia et al., 2002) it was found that the di-
vergence angle of consecutive leaves is similar for the
same cultivar and is approximately the Fibonacci an-
gle of 137.5
◦
. The handedness of the date palm is
the direction in which the leaves are born, clockwise
or counter clockwise. The frequencies of right or left
handedness in this study were more or less equal. Oil
palm measurements, testing for the vertical distance
between consecutive leaves, was measured in (Rees,
1964), showing that the horizontal distance between
leaves changes only gradually along the length of the
trunk.
The arrangement of the stubs of pruned leaves on
the trunk of the palm tree can be modelled accurately
by the cylindrical model. Arrangement of the leaves
on the tree crown, on the other hand, deviates from the
cylindrical model because the radius of the trunk on
the crown decreases. In this paper we used the cylin-
drical model with adaptations allowing for changes in
the trunk’s radius. This model is relatively accurate,
while maintaining simplicity and offering an analyti-
cal solution.
3.1 Mathematical Model Description
The mathematical model of the palm tree describes
both leaf growth locations and leaf growth angles,
on the trunk. Leaf growth locations are described
in cylindrical coordinates (θ, r, H) with respect to the
trunk, where θ is the angle, r is the radius, and H is
the height. Leaves are numbered according to their
age. The youngest leaf, situated at the top of the tree,
is marked as number 1 with consequent leaves being
marked according to the order of descent along the
tree trunk. The earlier the leaf budded - the higher the
number assigned to that specific leaf.
Being a leafing plant, the phyllotactic order of the
palm tree is not perfect, but still the location of the leaf
can be predicted quite accurately from the location
of its neighbour. Therefore, the phyllotactic model is
defined recursively. The location of leaf number n + i
can be calculated from the location of leaf n by the
Figure 3: Model of the “skeleton” of the palm tree created
using the mathematical model.
following equations:
θ
n+i
= θ
n
+ i · h · ψ (1)
r
n+i
= R
table
(n + i) · R (2)
H
n+i
= H
n
− i · d (3)
α
n+i
= α
table
(n + i) (4)
where:
• h - The handedness of the date palm, 1 for clock-
wise, -1 for counter clockwise.
• ψ - The Fibonacci angle of 137.5
◦
.
• R
table
( j) - A table with the ratio of trunk radius at
the growing point of leaf number j, to the radius
of the trunk at its widest point, R. The table was
measured from a reference tree assuming that this
value is representative.
• d - The vertical distance between two consecutive
leaves.
• α
n+i
- Leaf growing angle for leaf number n + i.
• α
table
( j) - A table with leaf growth angles for leaf
number j . The data was acquired from a reference
tree by measuring images taken from a perpendic-
ular angle to the leaf.
Only two parameters are unique for every tree, the
handedness of the tree h and the vertical distance be-
tween two consecutive leaves d. Knowing these pa-
rameters, the location of one leaf, and its age index,
VISAPP 2009 - International Conference on Computer Vision Theory and Applications
290
we can predict the locations of its neighbors. All val-
ues are computed proportional to the radius of the tree
whose value can not be recovered from the image.
Figure 3 shows a model of the palm tree created using
the aforementioned mathematical model.
The mathematical model describes a spiral phyl-
lotactic pattern, which can have different parastichy
numbers according to r and d. But still for most date
palm cultivars, including the one considered here, the
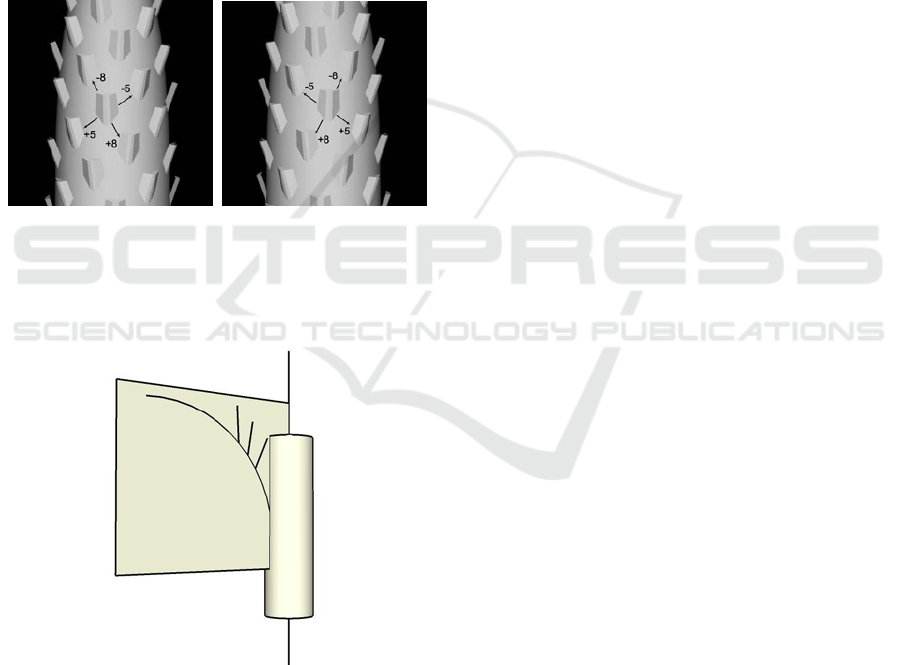
parastichy numbers are 5 and 8 (Figure 4).
Another assumption of our model is that the leaves
grow outward from the axis of the trunk. In other
words the leaf midribs are located on a plane con-
taining the trunk axis and the leaf growing point (see
Figure 5).
(a) (b)
Figure 4: Contact parastichy pattern for (a) right handed
tree and (b) a left handed tree.
Figure 5: Leaves grow outward from the axis of the trunk.
4 ALGORITHM DESCRIPTION
An overview of our proposed algorithm is summa-
rized above. The different stages are summarized in
the following subsections and the output of each stage
is illustrated in Figure 6. The input of the algorithm
is an image of a palm tree, Figure 6(a) is an example
of such an image.
• Calculating a per pixel probability image of the
leaves and trunk (Figure 6(b), Subsection 4.1).
• Searching for the location of the trunk and “tree
center” using the aforementioned images (Figure
6(c), Subsection 4.2).
• Creating, using the leaf probability image and the
“tree center”, a leaf clues image (Figure 6(d), Sub-
section 4.3).
• Search for prominent leaves using the leaf clues
image (Figure 6(e), Subsection 4.4).
• Estimation of the model parameters by using the
location of the prominent leaves and their growth
angles (Figure 6(f), Subsection 4.5).
• Search for more leaves using prediction based on
the model parameters (Figure 6(g), Subsection
4.6).
• 3D reconstruction of the palm tree is achieved by
use of the tree model parameters and the location
of the leaves on the 2D image (Figure 6(h), Sub-
section 4.7).
4.1 Calculating Probability Image of
the Leaves and Trunk
The interesting part of the leaf of the date palm is its
midrib - the central “spine” of the leaf. These midribs
and the trunk are the tree’s “skeleton”. The leaflets
can be added easily to this “skeleton”, creating a com-
plete model of the tree. The prominent feature of the
leaves is their green color, while the distinctive fea-
ture of the midrib is its smoothness - low level of gra-
dient. Generating the leaf midrib probability image -
P
l
, denoted as the “leaf probability image”, is based
on simple histogram learning techniques described in
(Jones and Rehg, 2002). Midrib and non-midrib his-
tograms were collected using a training set of manu-
ally labelled images. Given these histograms a four
dimensional table with the probability that a given
color and gradient defines the leafs’ midrib is calcu-
lated (see Figure 7). The table is applied to the origi-
nal image to get the leaf probability image. The size
of the histogram is 30 bins/channel for the HSV chan-
nels, and 10 bins/channel for the gradient. In practice,
USING PHYLLOTAXIS FOR DATE PALM TREE 3D RECONSTRUCTION FROM A SINGLE IMAGE
291

(a) Original image. (b) leaf and trunk proba-
bility images.
(c) Trunk location and
“tree center”.
(d) Leaf clues image.
(e) Prominent leaves
search.
(f) Model prediction. (g) Search for more
leaves.
(h) VRML 3D model.
Figure 6: Output of algorithm stages. (e)-(g) show a zoomed in section of the original image.
the combination of color and smoothness has proved
to be both informative and robust.
The dominant feature of the trunk is its color. The
trunk probability image - P
t
is calculated in the same
way as the leaf probability image only without con-
sidering the gradient. The leaf and trunk probability
images are illustrated in Figure 6(b) in green and blue
respectively.
4.2 Search for Trunk Location and
“Tree Center”
The location of the trunk is represented by three pa-
rameters: the horizontal location on the image in pix-
els - x, trunk radius in pixels - r, and the leaning angle
of the trunk toward the camera - γ (see Figure 8(a)).
Assuming that the image was taken nearly parallel to
the horizon, and that the palm trunk is perpendicular
to the ground, knowing these three parameters, en-
ables the calculation of the 3D location of the trunk,
and the transformation between location on the 3D
trunk to the image and vice versa. We define the tree
center as the point on the line of symmetry of the
trunk, with the vertical location on the image in pix-
els - y, immediately beneath the oldest leaf (see Figure
Figure 7: 3D “slice” of the 4D leaf probability table, defines
the probability of a given color with gradient size: 6-8. The
probability is illustrated by a ball whose size is proportional
to the probability.
8(a)). We call it the tree center because it is almost the
intersection point of all the leaves midribs line. The
tree center will be used later for the removal of leaf
VISAPP 2009 - International Conference on Computer Vision Theory and Applications
292

clues not in the direction “intersecting” with the tree
center.
Estimation of the four trunk location and tree cen-
ter parameters (x, r, y, γ) is done by searching for the
best location of a schematic model of the tree on both
the trunk and the leaf probability images. The model
is illustrated in Figure 8(b). Assuming that γ is not
too large, the trunk can be represented by a vertical
rectangle on the image plane. The “trunk rectangle”
is separated into two rectangles, D - descending and
U - ascending from the common “tree center”. Rect-
angles R and L represent the outer border of the trunk,
an area that should have pixels with low probability
in the trunk probability image. A leafy area is repre-
sented by rectangle M which should have a high prob-
ability for pixels in the leaf probability image. The
location and size of these rectangles are functions of
x, r and y.
For performance reasons the search is done in
phases. The first phase searches horizontally for x
and r, using the “Integral Image” algorithm (Viola and
Jones, 2001) it maximizes the energy function:
E
h
(x, r) = ( f (U ∪ D, P
t
) − f (L , P
t
) − f (R, P
t
)) · r
1
3
,
(5)
where f (A, P) =
∑
(i, j)∈A
i, j
P(i, j)
k
A
k
represents the average
probability of the pixels in the rectangle A on the
probability image P. The term r
1
3
was added to pre-
vent very thin trunks from being accepted. For every
local maximum a search for y is done using pattern
search (Lewis and Torczon, 2000) by maximizing:
E
v
(x, r, y) = ( f (M, P
l
) + f (D, P
t
) − f (U, P
t
)). (6)
The best (x, r, y) combination is chosen by maximiz-
ing:
E(x, r, y) = E
v
(x, r) · E
h
(x, r, y). (7)
Finally the best γ is chosen. Both left and right trunk
line-boundary angles are searched for in order to max-
imize the derivative over the trunk probability image.
The intersection point of these lines is used in order
to calculate γ. Results of the search for trunk loca-
tion and tree center are illustrated in Figure 6(c). The
tree center is marked by a white cross, and the trunk
borders are drawn in magenta.
4.3 Creating the Leaf Clues Image
The leaf clues image is created from the leaf proba-
bility image - P
l
by thresholding. After thresholding,
groups of connected pixels that we call leaf clues re-
main. The dominant clue direction is calculated using
PCA. Clues whose extensions do not intersect with
a circle around the tree center (whose radius is
3
2
· r)
are removed. Figure 6(d) shows an example of a leaf
clues image, with the tree center marked.
(a) (b)
Figure 8: (a) Trunk location and “Tree Center” parameters
description. (b) Rectangles U, D, R, L, and M, used in the
search for the trunk location and “Tree Center”.
4.4 Search for Prominent Leaves
The purpose of prominent leaf search is to find
enough leaves to be able to calculate the model pa-
rameters. Figure 6(e) illustrates the result of this
search. The leaf search is done on the leaf clues im-
age from bottom up because the lower leaves (older)
always hide the younger leaves above them. Only the
frontal leaves are considered because they are more
prominent and distinct. The process is iterative and
includes three parts: locating the leaf’s start point in
the leaf clues image, tracking this leaf from its start
point, and removing the leaf found from the leaf clues
image and the leaf probability image. These parts are
explained below.
Locating the Leaf Start Point: The leaf start point
is chosen as the lower point of a clue in the leaf clues
image. The clue is chosen from a region in the center
of the trunk, above the tree center.
Tracking the Leaf: The leaf is tracked from its
growing point using the leaf probability image and
the original image in HSV color space. Tracking is
done using segments whose length is one third of the
trunk’s radius. The end of each segment is the be-
ginning of the following segment but in a different
direction (Figure 9). The width and length of every
segment is calculated according to the model and the
leaf starting point. The tracking is done using a par-
ticle filter (Arulampalam et al., 2002), with a weight
function which is the product of the following terms:
• Sum of leaf pixels on the leaf probability image
plus the number of pixels that belong to this spe-
cific clue in the proposed leaf. This encourages
the leaf model to be on the leaf pixels and on the
starting clue (illustrated in green in Figure 9).
• 1/sum of the standard deviations of the pixels
USING PHYLLOTAXIS FOR DATE PALM TREE 3D RECONSTRUCTION FROM A SINGLE IMAGE
293
along the lines of the leaf segments (illustrated by
the blue lines in Figure 9). This discourages cross-
ing over to other leaves because lines along a leaf
tend to be smooth.
• Number of pixels of the proposed leaf that are not
sky pixels. A simple definition for a sky pixel in
HSV color space was determined empirically.
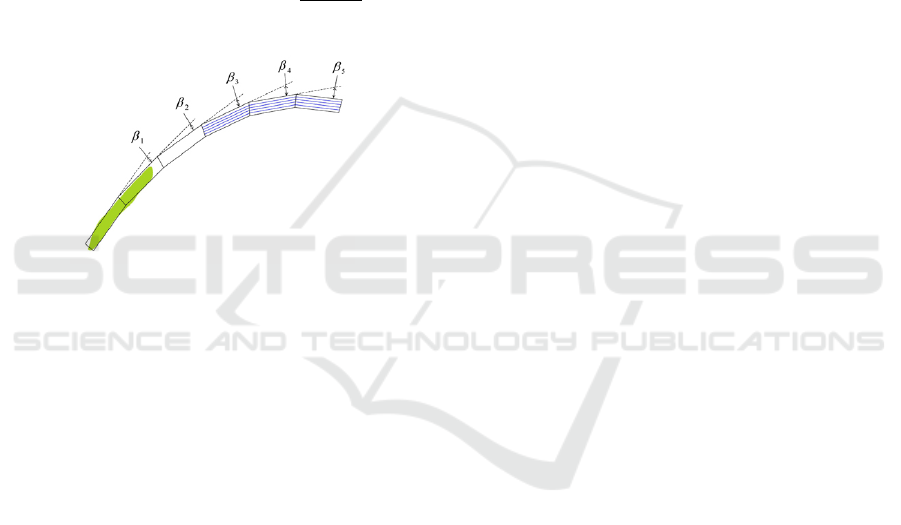
• 1/sum of the ascending angles. Consider the an-
gles between successive leaf parts, illustrated in
Figure 9 as β
1
−β
5
. Ascending angles are defined
as negative angles. They are counted to discour-
age the leaf from ascending against gravity.
Leaf samples are propagated by adding segments
with the mean direction of the previous segment per-
tubrated by Gaussian noise with σ =
leaf radius
4
relative
to the end of the segment.
Figure 9: Particle filter tracking model.
Removing the Leaf: Once the geometric shape of
the leaf has been found, it is removed from leaf clues
image and the leaf probability image.
4.5 Estimating Model Parameters
The exact location of neighboring leaves depends
on the leaf parameters and the tree model equa-
tions. Nevertheless, parameter similarities between
trees give similar parastichy patterns, making it possi-
ble to estimate the location of a neighboring leaf while
accounting for the handedness (possible locations are
calculated by using a range of possible parameters).
The first step in model parameter estimation is
done twice; once for the possibility of right hand-
edness and once for left handedness. Leaves found
in the last step are matched into a list of neighboring
couples according to their relative location. d is cal-
culated for every couple from the location of the two
leaves according to Equation 3. From the list of the
d’s outliers are removed and the handedness of the
tree is chosen to be the one with the most inliers and
d is computed as the mean value of the inliers.
The relative age index between leaf couples is
known according to their relative location and hand-
edness. All leaves are numbered according to their
age index in relation to the lowest leaf. The leaf index
of the lowest leaf is chosen as the one with the mini-
mal sum of differences between the leaf angle as seen
in the image in relation to the prediction of the leafs’
angle according to the model using the projection of
α from Equation 4 on to the image.
Figure 6(f) is the tree image. Prominent leaves are
marked in green with their age indexes, while those
marked in magenta predict the location and angle of
other leaves according to the model.
4.6 Search for More Leaves
Model parameters and prominent leaf age indexes en-
able the prediction of neighboring leaf locations and
growth angles using the tree model equations. The
search for more leaves is done iteratively by search-
ing for neighboring leaves from the bottom up. Every
tree location and growth angle of neighboring leaves
is calculated. If a leaf clue exists in the leaf clue im-
age in that area, then leaf tracking is performed in the
same way as in Section 4.4. After the search a leaf
is accepted if the percentage of pixels in the geomet-
ric shape of the leaf, found in the leaf clue image, is
beyond a given threshold. Figure 6(g) illustrates the
result of this search. All the leaves found, including
the ones found in the search for the prominent leaves
are marked in magenta.
4.7 3D Reconstruction
As mentioned in Section 3.1, one of the assumptions
of the tree model is that the leaves grow outward from
the axis of the trunk. Using this assumption, knowing
the location of the trunk and the location of the leaf
midrib on the 2D image and including the leaf grow-
ing point, we are able to calculate the location of the
3D midrib. Figure 6(h) is a 3D VRML model of the
date palm tree based on these calculations.
5 EXPERIMENTS
The algorithm described in this paper has been imple-
mented in MATLAB and tested on nearly 50 images
of date palm trees. In the first stage the leaf, trunk and
sky probability image were generated from a small
number of manually labeled images. The radius table
R
table
( j) and the leaf angle table α
table
( j) were mea-
sured from a small set of images of a tree.
Several results of the execution of the algorithm
are presented in Figure 10. Additional results as well
as the VRML models can be viewed at:
http://mis.hevra.haifa.ac.il/∼ishimshoni/Phyllotaxis/.
VISAPP 2009 - International Conference on Computer Vision Theory and Applications
294

6 CONCLUSIONS AND FUTURE
WORK
In this paper, we proposed a new 3D reconstruction
algorithm which reconstructs the 3D model of the
date palm tree from a single uncalibrated input image
by using the spiral phyllotactic pattern. To demon-
strate the effectiveness of the proposed method, we
show several output VRML model images.
3D reconstruction from a single input image is,
naturally, only the first step in the development of au-
tomatic vision algorithms to guide autonomous palm
tree maintenance machines. We envision the next step
to be the further development of the algorithm by in-
corporating structure from motion techniques in or-
der to improve the accuracy of the algorithm and to
integrate the algorithm into a robotic prototype. The
proposed algorithm can also be applied to additional
phyllotactic plants.
REFERENCES
Adler, I., Barabe, D., and Jean, R. (1997). A history of the
study of phyllotaxis. Ann. of Botany, 80(3):231–244.
Aracil, R., Saltar
´
en, R., and Sabater, J. (1999). Trepa: Par-
allel climbing robot for maintenance of palm trees and
large structures. In Proc. of Int. Workshop and Conf.
on Climbing and Walking Robots, pages 453–461.
Arulampalam, M. S., Maskell, S., Gordon, N., and Clapp,
T. (2002). A tutorial on particle filters for on-
line nonlinear/non-gaussian bayesian tracking. IEEE
Transactions on Signal Processing, 50:174–188.
Atela, P. and Gol
´
e, C. (2008). Phyllotaxis - an interactive
site for the mathematical study of plant pattern forma-
tion. www.math.smith.edu/∼phyllo/.
Cummings, F. W. and Strickland, J. C. (1998). A model of
phyllotaxis. J. of Theo. Biology, 192(4):531–544.
Elhoumaizia, M. A., Lecoustreb, R., and Oihabic, A.
(2002). Phyllotaxis and handedness in date palm
(phoenix dactylifera l.). Fruits, 57(5-6):297–303.
Ferry, M. (1998). The phyllotaxis of the date palm (phoenix
dactylifera l.). In Proc. Inter. Conf. on Date Palms, Al-
Ain, UAE, pages 559–571.
Fowler, D. R., Hanan, J., and Prusinkiewicz, P. (1989).
Modelling spiral phyllotaxis. Computers and Graph-
ics, 13(3):291–296.
Fowler, D. R., Prusinkiewicz, P., and Battjes, J. (1992). A
collision-based model of spiral phyllotaxis. In Proc.
ACM Conf. on Computer Graphics and Interactive
Techniques, pages 361–368.
Haering, N. C. and da Vitoria Lobo, N. (1999). Features
and classification methods to locate deciduous trees in
images. Comp. Vis. and Im. Under., 75(1-2):133–149.
Jean, R. V. (1994). Phyllotaxis: a systemic study in plant
morphogenesis. Cambridge University Press.
Jones, M. J. and Rehg, J. M. (2002). Statistical color mod-
els with application to skin detection. International
Journal of Computer Vision, 46(1):81–96.
Kaewapichai, W., Kaewtrakulpong, P., Prateepasen, A., and
Khongkraphan, K. (2007). Fitting a pineapple model
for automatic maturity grading. In ICIP, pages I: 257–
260.
Lewis, R. M. and Torczon, V. (2000). Pattern search algo-
rithms for linearly constrained minimization. SIAM
Journal on Optimization, 10(3):917–941.
Lintermann, B. and Deussen, O. (1999). Interactive model-
ing of plants. IEEE Computer Graphics and Applica-
tions, 19(1):56–65.
Martinez, A. R., Mart
´
ın, I., and Drettakis, G. (2004).
Volumetric reconstruction and interactive rendering
of trees from photographs. ACM Trans. Graph,
23(3):720–727.
Prusinkiewicz, P. and Lindenmayer, A. (1990). The Algo-
rithmic Beauty of Plants. Springer-Verlag.
Rees, A. R. (1964). The apical organization and phyllotaxis
of the oil palm). Annals of Botany, 28:57–69.
Reinhardt, D. (2005). Regulation of phyllotaxis.
International Journal of Developmental Biology,
49(5/6):539–546.
Ripin, Z. M., Soon, T. B., Abdullah, A. B., and Samad, Z.
(2000). development of a “low cost” modular pole
climbing robot. TENCON, 1:196–200.
Rutishauser, R. (1988). Symmetry in Plants, chapter Plas-
tochron ratio and leaf arc as parameters of a quantita-
tive phyllotaxis analysis in vascular plants, pages 171–
212. World Scientific Publications.
Shamsi, M. (1998). Design and development of a tree
climbing date harvesting machine. PhD thesis, Silsoe
College, Cranfield University, UK.
Shlyakhter, I., Rozenoer, M., Dorsey, J., and Teller, S.
(2001). Reconstructing 3D tree models from instru-
mented photographs. IEEE Computer Graphics and
Applications, 21(3):53–61.
Smith, R. S., Guyomarc’h, S., Mandel, T., Reinhardt, D.,
Kuhlemeier, C., and Prusinkiewicz, P. (2006). A plau-
sible model of phyllotaxis. Proceeding - National
Academy of Sciences USA, 103(5):1301–1306.
Steeves, T. A. and Sussex, I. M. (1989). Patterns in plant
development. Cambridge University Press.
Teng, C. H., Chen, Y. S., and Hsu, W. H. (2006). An
approach to extracting trunk from an image. IEICE
Transactions, E89-D(4):1596–1600.
Viola, P. and Jones, M. (2001). Rapid object detection using
a boosted cascade of simple features. In Proceedings
CVPR, pages I:511–518.
USING PHYLLOTAXIS FOR DATE PALM TREE 3D RECONSTRUCTION FROM A SINGLE IMAGE
295

Original images Segmented leaves VRML 3D model
Figure 10: Results of the algorithm’s operation.
VISAPP 2009 - International Conference on Computer Vision Theory and Applications
296
