
A STUDY OF GENETIC PROGRAMMING VARIABLE
POPULATION SIZE FOR DYNAMIC OPTIMIZATION PROBLEMS
Leonardo Vanneschi
Department of Informatics, Systems and Communication (D.I.S.Co.), University of Milano-Bicocca, Milan, Italy
Giuseppe Cuccu
Istituto Dalle Molle di Studi sull’Intelligenza Artificiale (IDSIA), Lugano, Switzerland
Keywords:
Genetic programming, Variable size population, Dynamic optimization.
Abstract:
A new model of Genetic Programming with variable size population is presented in this paper and applied to
the reconstruction of target functions in dynamic environments (i.e. problems where target functions change
with time). The suitability of this model is tested on a set of benchmarks based on some well known symbolic
regression problems. Experimental results confirm that our variable size population model finds solutions of
similar quality to the ones found by standard Genetic Programming, but with a smaller amount of computa-
tional effort.
1 INTRODUCTION
Many real-world problems are anchored in dynamic
environments, where some element of the problem
domain, typically the target, changes with time. For
this reason, developing solid evolutionary algorithms
(EAs) to reliably solve these problems is an important
task. In the last few years, many contributions have
appeared which studied dynamic optimization envi-
ronments and developped new evolutionary frame-
works for solving them. Nonetheless, the majority
of those approaches are based on Genetic Algorithms
(GAs) (Goldberg, 1989) or Particle Swarm Optimiza-
tion (PSO) (Clerc, 2006) and the problem objective is
to find the extrema (maxima or minima) of a target
function that changes with time. On the other hand,
very few contributions have appeared to date that
study the ability of Genetic Programming (GP)(Koza,
1992) to reconstruct target functions on dynamic op-
timization environments.
In this paper we hypothesize that variable size
population GP is a promising method for dynamic op-
timization problems. This idea is not new in evolu-
tionary computation; for instance, it has been applied
to PSO in (Fernandes et al., 2005). However, it has
never been applied to GP before. We propose a vari-
able size population GP model called DynPopGP. It
is inspired by the one presented in (Tomassini et al.,
2004). Simply speaking, it works by shrinking the
population when fitness is improving and increasing
its size, by adding new genetic material, when the
evolution stagnates. Our hypothesis is that when the
target function changes, evolution of the current pop-
ulation should stagnate. Thus the evolution should
benefit from the creation of new genetic material, that
should give the necessary amount of diversity to start
the optimization of a new target function.
This paper is structured as follows: in Section 2
we discuss previous contributions in dynamic opti-
mization. In Section 3 we discuss the reasons why
it is not suitable to directly apply the GP model pre-
sented in (Tomassini et al., 2004) to dynamic opti-
mization and we present DynPopGP that extends it.
Section 4 contains a description of the test problems
and presents the experimental setting used in this pa-
per. In Section 5 we show the obtained experimental
results. Finally, Section 6 concludes the paper and
suggests directions for future research.
2 DYNAMIC OPTIMIZATION
Over the past few years, a number of authors have ad-
dressed the problem of EAs premature loss of diver-
sity in dynamic environments in many different ways.
119
Vanneschi L. and Cuccu G. (2009).
A STUDY OF GENETIC PROGRAMMING VARIABLE POPULATION SIZE FOR DYNAMIC OPTIMIZATION PROBLEMS.
In Proceedings of the International Joint Conference on Computational Intelligence, pages 119-126
DOI: 10.5220/0002314701190126
Copyright
c
SciTePress

Surveys of these studies can be found for instance
in (Branke, 2001; Branke, 2003). More recent con-
tributions include (Yang, 2004) where, based on the
concept of problem difficulty, a new dynamic envi-
ronment generator using a decomposable trap func-
tion is proposed; (Huang and Rocha, 2005), where
a coevolutionary agent-based model is used; (Rand
and Riolo, 2005) where the use of mutation for diver-
sity maintenance is investigated and (de Franc¸a et al.,
2005), where the use of artificial immune networks
for multimodal function optimization on dynamic en-
vironments is studied.
All the above quoted contributions treat the prob-
lem of tracking the extrema in a dynamic envi-
ronment, where the target function changes with
time and concern GAs, PSO or other EAs variants.
Very few contributions have appeared to date deal-
ing with the (more complex) problem of approximat-
ing/reconstructing target functions that change with
time by means of GP. Noteworthy recent exceptions
are: (Dempsey, 2007), where financial time series (in-
dex closing price data) are reconstructed by means
of Grammatical Evolution, and (Tanev, 2007) where
an approach for incorporating learning probabilistic
context-sensitive grammar in GP is employed for the
evolution and adaptation of locomotion gaits of a sim-
ulated snake-like robot. Nevertheless, both these ap-
proaches use Grammar-Based GP and employ it for
very particular and complex applications.
The goal of this paper is different: first of all, we
want to study standard tree-based GP (Koza, 1992),
and one variant thereof using variable size popula-
tions; secondly, we want to present and employ (here
for the first time) more simple, and thus easier to
study, test problems. The proposed GP framework is
presented in Section 3 and the used test functions in
Section 4.
3 VARIABLE SIZE POPULATION
GP
In 2003, an idea for counteracting the negative ef-
fects of bloat (Banzhaf and Langdon, 2002; Poli et al.,
2008) and of premature convergence (Burke et al.,
2002) on GP was presented. It consisted in reduc-
ing the size of populations at a linear rate (Fern´andez
et al., 2003c; Fern´andez et al., 2003b). This was
achieved by removing a fixed number of individuals
at each generation. This technique was called plague
and it has been shown to have some positive effects
on GP systems. That idea started from the observa-
tion of a general behavior of GP over a wide set of
problems: normally fitness improves quickly at the
beginning of GP runs and, after a number of gen-
erations, improvements are more difficult to obtain.
In this second phase, plagues allow to save computa-
tional effort, that would be wasted otherwise, since it
does not bring appreciable advantages.
On the other hand, it is clear that even if a con-
siderable amount of computational effort is saved, the
blind deletion of individuals at each generation prob-
ably cannot lead to the discovery of better individuals
than the ones found by the standard GP process. Fur-
thermore, steadily decreasing populations produce a
progressive loss of diversity, especially at the geno-
typic level. For this reason, in (Tomassini et al.,
2004), an extension of the plague technique aimed
at varying the population size in an intelligent way
during the execution of each GP run, was presented.
In that model, adds and suppressions of individuals
are operated dynamically on the basis of the behavior
of the GP system: population size is decreased while
the algorithm is progressing (i.e. fitness is improv-
ing) and it is increased when the algorithm reaches
the stagnation phase. In this way, when the algorithm
is progressing, as much computational effort as pos-
sible is saved and this previously saved effort is spent
only when it is really useful, i.e. when the algorithm
is stagnating and new genetic material is needed.
In (Tomassini et al., 2004) the decision whether to
shrink or inflate the population was taken on the ba-
sis of the relationship between the best fitness value
in the population at the current generation g (bf
g
) and
the one at the previous generation (bf
g−1
). This value
was stored in a variable the authors called pivot. Two
versions of pivot are presented in (Tomassini et al.,
2004): in the first one pivot= ∆
g−1
/∆
g
and in the sec-
ond one pivot= ∆
g−1
− ∆
g
, where ∆
g
= bf
g−1
− b f
g
.
The GP model using the first version of pivot was
called DIV, while the one using the second version
was called SUP in (Tomassini et al., 2004).
In Section 3.1 we discuss the reasons why DIV
and SUP are not suitable to solve dynamic optimiza-
tion problems and in Section 3.2 we present our
new variable size population GP model, called Dyn-
PopGP, that extends DIV and SUP.
3.1 DIV and SUP in Dynamic
Environments
Both the DIV and SUP methods introduced
in (Tomassini et al., 2004) have the following
characteristics:
(i) The decision on whether to shrink or inflate the
population is taken only on the basis of the re-
lationship between the best fitness values at the
current generation and at the previous one. This
IJCCI 2009 - International Joint Conference on Computational Intelligence
120

decision does not depend on how good those fit-
ness values are. In other words, this decision is
the same independently from the fact that GP has
found good solutions or bad ones.
(ii) The quantity of individuals that have to be added
to or suppressed from the population depends on
the current population size (in both DIV and SUP
when individuals have to be suppressed, 1% of the
populationis suppressed and when they need to be
added, a number of individuals equal to the 0.2%
of the population is added). Thus, additions and
suppressions are more violent when the popula-
tion is large.
A consequence of point (i) is that, when applied to
dynamic optimization, DIV and SUP behave exactly
the same in case the algorithm stagnates on a particu-
lar target function (but the target function remains the
same) and in case the target function changes. How-
ever, when the target function changes, and in partic-
ular if the new target function is “different enough”
from the old one, we expect a more violent worsen-
ing in fitness than when the algorithm stagnates on a
fixed target function. In particular, if we use an elitist
algorithm (i.e. we copy the best, or a pool of good
individuals, unchanged in the next population at each
generation), the best fitness in the population cannot
worsen if the target function stays the same. It can
only worsen if the target function has changed (be-
cause the best individual at the previous generation
may change its fitness). The algorithm we propose
(DynPopGP) behaves in two different ways in these
two different situations.
A consequence of point (ii) is that, when the target
function changes, the population size may grow indef-
initely. In fact, suppose DIV or SUP are optimizing
a given target function and are in a stagnation phase.
Then, the population keeps growing. Now, assume
the target function changes. The population will con-
tinue growing up until the new genetic material nec-
essary to optimize the new target function has been
created. DynPopGP solves this problem by defining
a new function to quantify the amount of individuals
that have to be added or deleted from a population.
For these reasons, we do not consider the DIV
and SUP models any longer in this paper. A detailed
experimentation showing the practical advantages of
DynPopGP compared to DIV and SUP is definitely
needed in the future.
3.2 New Variable Size Population Model
The DynPopGP algorithm we propose can be sum-
marized by the pseudo-code in Figure 1, where
we consider minimization problems (i.e. small fit-
ness values are better than large ones)
1
. This
algorithm uses a number of parameters (trg fit,
old got trg, new got trg, stand by size) and functions
(update pop size, ∆
pop
) that we describe below. Em-
pirical values for these parameters, coming from a set
of preliminary experiments, have been used in this
work. More detailed sensitivity analysis on these pa-
rameters definitely deserves to be conducted in the fu-
ture.
begin
Generate a population of N random individuals;
best = best individual in the population;
old got trg = false;
for g := 1 to maxgen do
new got trg = (fitness(best) ≤ trg fit);
if (not new got trg)
then
elitism (i.e. copy of the best);
selection;
reproduction / crossover;
mutation;
best = best individual in the new population;
new got trg = (fitness(best) ≤ trg fit);
if (old got trg)
then
// The old best had reached the target, while
// the new best has not reached it:
// the target function has surely changed.
// Set the population size to the initial size
update pop size(N - current pop size);
else
// Neither the new best, nor the old best
// have reached the target: update the
// population size using the ∆
pop
function
update pop size(∆
pop
());
endif
else
if (not old got trg)
then
// The new best has reached the target, while
// the old best had not reached it. This means
// that the target has been found now.
// I have to spend as few computational effort
// as possible until the target function changes
// (or the process terminates).
// I set the population size to a prefixed
// “stand-by” value
update pop size(stand by size - current pop size);
endif
endif
old got trg = new got trg;
endfor
end
Figure 1: Pseudo-code for the DynPopGP algorithm.
1
The authors are aware that in case of minimization
problems the term fitness is rather incorrect. Nevertheless,
they keep using it for simplicity.
A STUDY OF GENETIC PROGRAMMING VARIABLE POPULATION SIZE FOR DYNAMIC OPTIMIZATION
PROBLEMS
121

trg fit represents a fitness value (target) that ap-
proximates the optimum. In this work, we have used
a value equal to 0.01.
old got trg (respectivelynew got trg) is a boolean
variable whose value is true if the best fitness in the
population at the previous (respectively current) gen-
eration is better than or equal to the target and false
otherwise.
stand by size is a small value of the population
size that is used when the optimum of a target func-
tion has been approximated in a satisfaisable way, and
we have to wait for the target function to change. In
this case, the population has to be as small as pos-
sible, so that we can save computational effort. In
our work, we wanted to set this value as a function
of the initial population size. We have chosen a value
of stand by size equal to the initial population size di-
vided by 4 because, by means of a set of experiments,
we have seen that this value represents a good com-
promise between saving computational effort (popu-
lation shrinking) and keeping some good genetic ma-
terial in the population.
update pop size(x) is a function that adds |x| in-
dividuals to the population if x is positive and sup-
presses |x| individuals from the population if x is neg-
ative. When |x| individuals have to be suppressed, the
population is sorted. The 2 · x worst (in terms of fit-
ness) individuals in the population are consideredand,
among these individuals, the x largest ones (in terms
of number of tree nodes) are suppressed (as it hap-
pened for the DIV and SUP algorithms). When |x|
individuals have to be added, they are randomly gen-
erated with the same initialization method that is used
at the beginning of the GP run (ramped half-and-half
in this work).
∆
pop
() is a function that returns the number of in-
dividuals that have to be to be added or suppressed
from the population when neither the old best fitness
value nor the new one approximate the optimal fit-
ness value in a satisfaisable way. We want ∆
pop
() to
be a function of the current best fitness in the popu-
lation and the current population size, which are the
two basic principles that were not taken into account
by the DIV and SUP algorithms. For doing this,
we define two new functions: best fit contribution
and pop size contribution and we multiply their re-
turned values. We want the result of this multiplica-
tion to be immediately interpretable by a human, so
we impose that the results of best fit contribution and
pop size contribution belong to the range [1, 10]. In
this way, their product belongs to the range [1, 100]
and it can be interpreted, for instance, as a percentage
(which represents the respective contributions given
by the best fitness value and the population size). The
∆
pop
() function performs the following calculation:
∆
pop
() = pivot· strength·best fit contribution() ·
pop size contribution()
where:
pivot is a variable whose value is −1 if the best
fitness in the population at the current generation is
better then the one at the previous generation and +1
otherwise (in practice, the value of pivot determines
if individuals have to be added or suppressed).
strength is a variable that determines how strong
populations inflate and deflate have to be at each
step, and it is used to rescale the value of
best fit contribution() · pop size contribution(). In
this work, we use a value equal to 0.3. In this way, the
maximum number of individuals that can be added to
or suppressed from the population is 30 (giventhat the
maximum possible value of best fit contribution() ·
pop size contribution() is 100). In fact, experimental
evidence confirms that adding more than 30 individu-
als at a time to the population eccessively increments
the computational effort without a correspondinggain
in the quality of the generated solutions.
The best fit contribution() function determines
the contribution given to the ∆
pop
() by the best fitness
value reached. As we have said above, we want this
function to return a value in the range [1, 10]. Further-
more, we want it to return 10 (maximum contribution
to the ∆
pop
()) when the best fitness in the population
is bad (fitness above a certain threshold, 60 in this
work) and to return the minimum value when the best
fitness in the population approximates the optimum in
a satisfaisable way. The easier way to obtain this, is to
define the best fit contribution() as a linear function,
for instance a straight line, that intersects the points
(trg fit,min coeff) and (max fit,max coeff) where
min coeff is equal to 1, max coeff is equal to 10 and
max fit is equal to 60. So, the best fit contribution()
function is defined by the pseudo-code in Figure 2.
best fit contribution() ::
if ( fitness(best) ≤ trg fit) then return min coeff;
elsif (fitness(best) ≥ max fit) then return max coeff;
else return
max coeff − min coeff ·
fitness(best)−trg fit
max fit−trg fit
+ min coeff
endif
Figure 2: Pseudo-code for the best fit contribution func-
tion.
Finally, the pop size contribution() function de-
termines the contribution to the ∆
pop
() given by
the current population size. Analogously to the
best fit contribution, we have used a linear function
that returns the maximum possible value (10 in this
work) when the current population size is minimal
(i.e. it is equal to stand by size, that has been set to
IJCCI 2009 - International Joint Conference on Computational Intelligence
122

the initial population size divided by 4 in this work)
and the minimum possible value (0 in this work) when
the current population size is maximal (i.e. smaller or
equal to stand by size).
4 TEST PROBLEMS AND
EXPERIMENTAL SETTING
Some benchmark problems have been defined for
testing the performances of optimization meth-
ods in dynamic environments. In particular,
Branke (Branke, 2001; Branke, 2003) defines and
uses moving peaks types of functions. In these bench-
marks, hand-tailored fitness landscapes are defined
and the positions of the extrema and their basins of at-
traction are modified with time. Similar problems are
also used, for instance, in (Huang and Rocha, 2005;
Rand and Riolo, 2005; de Franc¸a et al., 2005).
However, this type of benchmark is not suitable
for the present study. In fact, in this work, we want to
study the ability of GP to reconstruct dynamic target
functions and not follow moving extrema. With this
goal in mind, it would make no sense to use moving
peaks benchmarks as the ones presented in (Branke,
2001; Branke, 2003), given that, in those kinds of
benchmark, extrema are moved by changing some ad-
ditive or multiplicative constants to a (otherwise not
changing) target function. If one uses GP with linear
scaling (introduced in (Keijzer, 2003)), the moving
peaks problem reduces to a static GP problem, given
that linear scaling allows to reconstruct the shape of
the target functions, offering a method to automati-
cally determine additive and multiplicative constants.
For this reason, in this paper we define a new set
of benchmark problems that can be used to test GP
ability to reconstruct target functions in dynamic en-
vironments. These benchmarks are symbolic regres-
sion problems inspired by (Keijzer, 2003). In partic-
ular, maintaining the same terminology as in (Kei-
jzer, 2003), we have considered test functions F
12
,
F
13
, F
14
, F
15
and F
16
(presented in (Keijzer, 2003) at
page 9) and we have used them to build dynamic test
problems in which the importance of the modifica-
tion of the target function can be tuned. Even though
presented in (Keijzer, 2003), we also report here the
equations for these functions:
• F
12
(x,y) = xy + sin((x− 1)(y− 1))
• F
13
(x,y) = x
4
− x
3
+ y
2
/2− y
• F
14
(x,y) = 6 sin(x) cos(y)
• F
15
(x,y) = 8/(2+ x
2
+ y
2
)
• F
16
(x,y) = x
3
/5− y
3
/2− y− x
As in (Keijzer, 2003), for all these functions the
fitness cases are created by generating 20 random val-
ues (with uniform distribution) for x and y in the range
[−3,3].
We are aware that these test functions are bi-
dimensional and thus do not represent real-life ap-
plications (typically characterized by many features
and thus multi-dimensional), nevertheless, as reported
in (Keijzer, 2003) at page 8: “The aim of this set of
experiments is to demonstrate the practical implica-
tions of the use of the [method] studied here. Being of
low dimensionality does not make the problems easy
however. Many of the problems above mix trigonom-
etry with polynomials, or make the problems in other
ways highly non-linear”.
Using these test functions, we have built three
benchmarks for dynamic optimization that we have
called BENCH1, BENCH2 and BENCH3. The target
function at each generation is calculated by the algo-
rithm in Figure 3, where given a test function F
i
, with
12 ≤ i ≤ 15 succ(F
i
) = F
i+1
and succ(F
16
) = F
12
. The
begin
Define a set of test functions F = { f
1
, f
2
,..., f
n
}
for g := 1 to maxgen do
For each fitness case (x,y), the target value is:
n
∑
i=1
f
i
(x,y)
if (g mod period = 0) then
∀1 ≤ i ≤ n : f
i
:= succ( f
i
)
endif
endfor
end
Figure 3: Pseudo-code for target calculation in benchmark
problems BENCH1, BENCH2 and BENCH3 The differ-
ence between these benchmark is in the size of set F: n = 2
for BENCH1; n = 3 for BENCH2 and n = 4 for BENCH3.
difference between these benchmaks is in the cardi-
nality of the set of functions F used for calculating
the target: for BENCH1, F contains two functions.
These functions are F
12
and F
13
at generation 1. The
target value is calculated performing the sum of these
two functions for each couple of points (x, y). At each
period generations, one of the two functions changes
(i.e. it is deleted from set F and replaced by another
function), while the other stays the same, in a cyclic
way so that all the test functions are used.
BENCH2 is like BENCH1, except that F contains
3 functions, that are F
12
, F
13
and F
14
at generation 1
and at each period generations, one of them changes,
while the other two stay the same.
BENCH3 is similar, except that F contains 4 func-
tions, that are F
12
, F
13
, F
14
and F
15
at generation 1
and at each period generations, one of them changes,
while the other three stay the same.
In this way, BENCH1 has the more violent target
A STUDY OF GENETIC PROGRAMMING VARIABLE POPULATION SIZE FOR DYNAMIC OPTIMIZATION
PROBLEMS
123

modifications at each period generations, BENCH3
has the less violent modifications, while BENCH2 is
in an intermediary situation.
In this work, we have used a value of period equal
to 20. The other parameters used are as follows: pop-
ulation size of 200 individuals; function set equal to
{+,−,∗,/} (exactly the same method as in (Keijzer,
2003) has been used to avoid divisions with denomi-
nator equal to zero and thus to ensure operators clo-
sure); terminal set composed by two floating point
variables and four ephemeal random constants; max-
imum tree depth for initialization equal to 6; max-
imum tree depth for crossover and mutation equal
to 17; tournament size equal to 10; standard sub-
tree crossover (Koza, 1992) applied with probabil-
ity 0.9; standard subtree mutation (Koza, 1992) ap-
plied with probability 0.1; maximum number of gen-
erations equal to 100 (in this way, given that period=
20, the process stops when the target function returns
the same as at generation 1); generational GP with
elitism (i.e. copy of the best individual unchanged
in the next population at each generation). Fitness is
the root mean squared error (RMSE) between outputs
and targets. All the results reported in the next section
have been obtained by performing 100 independent
runs of each GP model (standard GP and DynPopGP)
for each banchmark. With standard GP we indicate
the canonic (fixed size population) GP process (Koza,
1992).
5 EXPERIMENTAL RESULTS
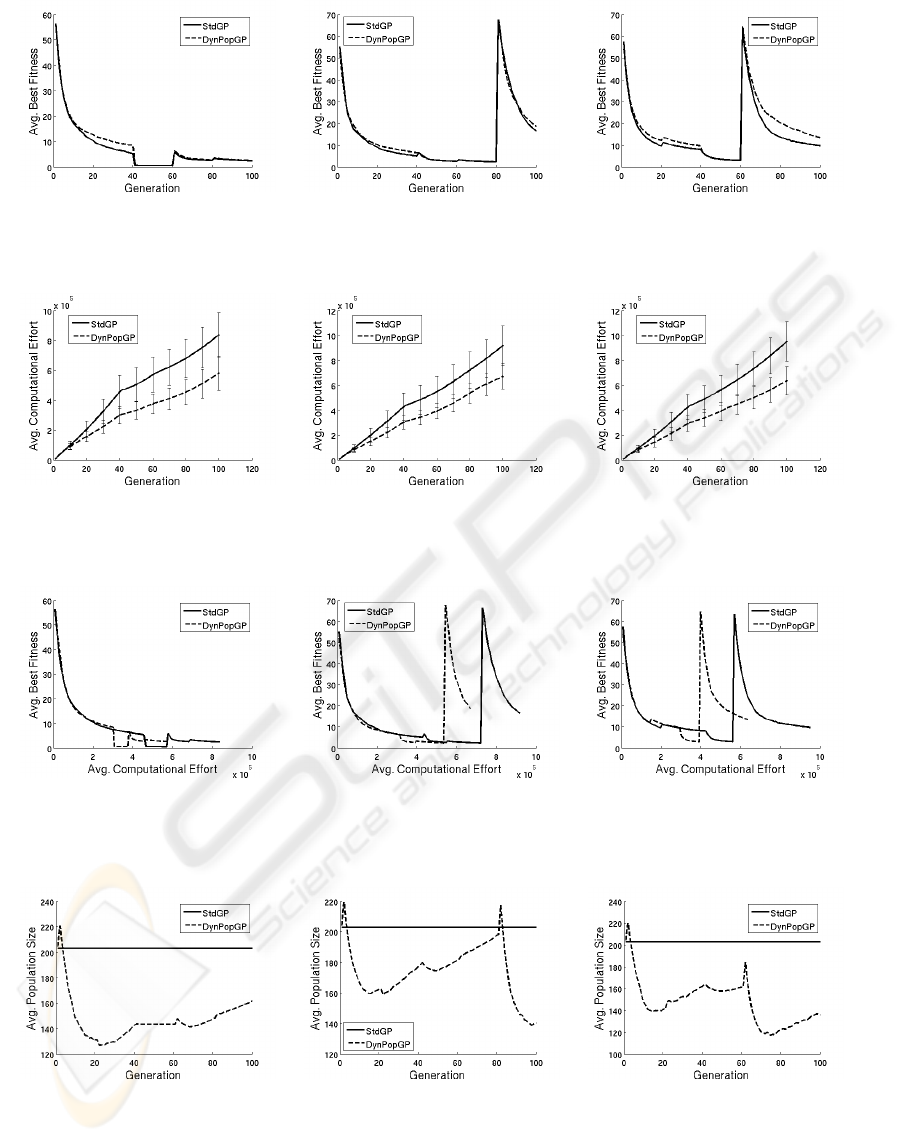
In Figure 4 we report average best fitness values over
100 independent runs against generations for stan-
dard GP (stdGP) and DynPopGP for BENCH1 (Fig-
ure 4(a)), BENCH2 (Figure 4(b)) and BENCH3 (Fig-
ure 4(c)). This figure clearly shows that the two GP
models find solutions of similar qualities at corre-
sponding generations for all the three studied bench-
marks (standard deviation error bars, not shown here
for simplicity, confirm that the differences between
the curves in Figure 4 are not statistically relevant).
Seen from this perspective, the two GP models might
seem equivalent. However, as reported for instance
in (Fern´andez et al., 2003a), comparing the perfor-
mances of two GP models against generations may
lead to wrong conclusions, given that GP individu-
als have a variable size and thus evaluating a genera-
tion for the two models may request a very different
amount of computational resources.
For this reason, in Figure 5 we report the values of
the computational effort against generations (values
averaged over the same 100 runs as in Figure 4). We
have considered exactly the same definition of com-
putational effort as in (Fern´andez et al., 2003a), i.e.
the computational effort at a given generation g (E
g
)
is given by: E
g
= PE
g
+ PE
g−1
+ ... + PE
1
, where the
partial effort at generation g (PE
g
) is defined as the
sum of the numbers of nodes of all the individuals in
the population at generation g. Given that fitness cal-
culation is often the most computationally expensive
part of an EA and that in GP this calculation largely
depends on the size of the individuals in the popula-
tion, this measure clearly gives an idea of the com-
putational complexity of executing a GP model (as
claimed in (Fern´andez et al., 2003a)). Figure 5 shows
that the effort spent by DynPopGP is smaller than the
one spent by stdGP for all the three studied bench-
marks. Standard deviations reported in figure as error
bars seem to hint that these results are statistically sig-
nificant.
Authors of (Fern´andez et al., 2003a) report results
of the average best fitness against computational ef-
fort. We do the same in Figure 6, where it is clear
that DynPopGP finds solutions of similar quality with
a smaller computational effort than stdGP.
In Figure 7 we report the average population size
at each generation (calculated using the same 100
runs as in the previous figures). We can see that
the population size of DynPopGP is always smaller
than the one of stdGP for all the three studied bench-
marks. Nonetheless, we can notice that the pop-
ulation size of DynPopGP, tends to grow at each
period generations (generation number multiple of
20), because of the modification in the target func-
tion. In some cases this growth begins slightly be-
fore the end of a period, probably because the partic-
ular target function had already been optimized and
the stagnation phase was beginning. Another inter-
esting thing to remark is that, after a first phase of
population shrinking, which is common in the three
benchmarks, the population growth is stronger for
BENCH1 (which has the more violent target modifi-
cations) than for BENCH3 (which has the less violent
target modifications), while the behavior of BENCH2
is intermediary. Furthermore, it is possible to see
that for BENCH1 the population size continues to
grow until the end of the run, while for BENCH2 and
BENCH3 there is a new phase in which the popula-
tion starts shrinking once again (at about generation
80 for BENCH2 and generation 60 for BENCH3).
6 CONCLUSIONS
This paper investigates the usefulness of variable size
population Genetic Programming (GP) on dynamic
IJCCI 2009 - International Joint Conference on Computational Intelligence
124
(a) (b) (c)
Figure 4: Average best fitness against generations for stdGP and DynPopGP. (a): BENCH1; (b): BENCH2; (c): BENCH3.
(a) (b) (c)
Figure 5: Computational effort against generations for stdGP and DynPopGP. (a): BENCH1; (b): BENCH2; (c): BENCH3;
(a) (b) (c)
Figure 6: Average best fitness against computational effort for stdGP and DynPopGP. (a): BENCH1; (b): BENCH2; (c):
BENCH3.
(a) (b) (c)
Figure 7: Population size against generations for stdGP and DynPopGP. (a): BENCH1; (b): BENCH2; (c): BENCH3.
problems. The idea is not new in Evolutionary Com-
putation (Fernandes et al., 2005), but, to the best of
our knowledge, it had never been extended to GP be-
fore. In particular, we believethat a model inspired by
the one presented in (Tomassini et al., 2004) should be
suitable for this kind of problems.
A STUDY OF GENETIC PROGRAMMING VARIABLE POPULATION SIZE FOR DYNAMIC OPTIMIZATION
PROBLEMS
125

Contributions of this paper are: first of all, we
have motivated the fact that the GP model presented
in (Tomassini et al., 2004), taken as it is, is not suit-
able for dynamic optimization problems. Succes-
sively, we have presented a GP model that extends
the one introduced in (Tomassini et al., 2004) and
that we candidate for suitably solving dynamic opti-
mizaton problems. We have called that model Dyn-
PopGP. We have also defined a new set of bench-
marks to test GP models for dynamic optimization,
based on some symbolic regression problems used
in (Keijzer, 2003). Finally, we have experimentally
shown that DynPopGP allows GP to save computa-
tional effort compared to standard GP, while finding
solutions of the same accuracy, at least for the studied
benchmarks.
This work is clearly a first and preliminary step in
this research track. The usefulness of GP (and in par-
ticular GP with variable size population) for dynamic
optimization deserves further investigation. In partic-
ular, GP models have to be tested on hard real-life ap-
plications, typically characterized by a large number
features and few samples and the issue of generaliza-
tion to out-of-sample data deserves to be investigated.
REFERENCES
Banzhaf, W. and Langdon, W. B. (2002). Some consider-
ations on the reason of bloat. Genetic Programming
and Evolvable Machines, 3:81–91.
Branke, J. (2001). Evolutionary Optimization in Dynamic
Environments. Kluwer.
Branke, J. (2003). Evolutionary approaches to dynamic op-
timization problems – introduction and recent trends.
In Branke, J., editor, GECCO Workshop on Evolution-
ary Algorithms for Dynamic Optimization Problems,
pages 2–4.
Burke, E., Gustafson, S., Kendall, G., and Krasnogor, N.
(2002). Advanced population diversity measures in
genetic programming. In J. J. Merelo et al., editor,
Parallel Problem Solving from Nature - PPSN VII,
volume 2439 of LNCS, pages 341–350. Springer.
Clerc, M. (2006). Particle Swarm Optimization. ISTE.
de Franc¸a, F. O., Zuben, F. J. V., and de Castro, L. N. (2005).
An artificial immune network for multimodal function
optimization on dynamic environments. In GECCO
’05: Proceedings of the 2005 conference on Genetic
and evolutionary computation, pages 289–296, New
York, NY, USA. ACM.
Dempsey, I. (2007). Grammatical Evolution in Dynamic
Environments. PhD thesis, University College Dublin,
Ireland.
Fernandes, C., Ramos, V., and Rosa, A. (2005). Varying the
population size of artificial foraging swarms on time
varying landscapes. In International Conference on
Artificial Neural Networks: Biological Inspirations,
volume 3696 of LNCS, pages 311–316. Springer.
Fern´andez, F., Tomassini, M., and Vanneschi, L. (2003a).
An empirical study of multipopulation genetic pro-
gramming. Genetic Programming and Evolvable Ma-
chines, 4(1):21–52.
Fern´andez, F., Tomassini, M., and Vanneschi, L. (2003b).
Saving computational effort in genetic programming
by means of plagues. In Congress on Evolutionary
Computation (CEC’03), pages 2042–2049, Canberra,
Australia. IEEE Press, Piscataway, NJ.
Fern´andez, F., Vanneschi, L., and Tomassini, M. (2003c).
The effect of plagues in genetic programming: A
study of variable size populations. In Ryan, C., et al.,
editor, Genetic Programming, 6th European Confer-
ence, EuroGP2003, Lecture Notes in Computer Sci-
ence, pages 317–326. Springer, Berlin, Heidelberg,
New York.
Goldberg, D. E. (1989). Genetic Algorithms in Search, Op-
timization and Machine Learning. Addison-Wesley.
Huang, C.-F. and Rocha, L. M. (2005). Tracking extrema in
dynamic environments using a coevolutionary agent-
based model of genotype edition. In GECCO ’05:
Proceedings of the 2005 conference on Genetic and
evolutionary computation, pages 545–552, New York,
NY, USA. ACM.
Keijzer, M. (2003). Improving symbolic regression with in-
terval arithmetic and linear scaling. In C. Ryan et al.,
editor, Genetic Programming, Proceedings of the 6th
European Conference, EuroGP 2003, volume 2610 of
LNCS, pages 71–83, Essex. Springer, Berlin, Heidel-
berg, New York.
Koza, J. R. (1992). Genetic Programming. The MIT Press,
Cambridge, Massachusetts.
Poli, R., Langdon, W. B., and McPhee, N. F. (2008). A
field guide to genetic programming. Published via
http://lulu.com and freely available at http://www.gp-
field-guide.org.uk. (With contributions by J. R. Koza).
Rand, W. and Riolo, R. (2005). The problem with a self-
adaptative mutation rate in some environments: a case
study using the shaky ladder hyperplane-defined func-
tions. In GECCO ’05: Proceedings of the 2005
conference on Genetic and evolutionary computation,
pages 1493–1500, New York, NY, USA. ACM.
Tanev, I. (2007). Genetic programming incorporating bi-
ased mutation for evolution and adaptation of snake-
bot. Genetic Programming and Evolvable Machines,
8(1):39–59.
Tomassini, M., Vanneschi, L., Cuendet, J., and Fern´andez,
F. (2004). A new technique for dynamic size popula-
tions in genetic programming. In Proceedings of the
2004 IEEE Congress on Evolutionary Computation
(CEC’04), pages 486–493, Portland, Oregon, USA.
IEEE Press, Piscataway, NJ.
Yang, S. (2004). Constructing dynamic test environ-
ments for genetic algorithms based on problem diffi-
culty. In Evolutionary Computation, 2004. CEC2004.
Congress on, volume 2, pages 1262–1269. IEEE, Pis-
cataway NJ, USA.
IJCCI 2009 - International Joint Conference on Computational Intelligence
126
