
PURE CO-EVOLUTION FOR SHAPE NESTING
Jeffrey Horn
Mathematics and Computer Science, Northern Michigan University
1401 Presque Isle Avenue, Marquette, Michigan, U.S.A.
Keywords: Shape nesting, Coevolution, Co-evolution, Cooperation, Cooperative Co-evolution, Fitness sharing,
Resource-defined fitness sharing, Speciation, Niching, Selection, Infinite population.
Abstract: We test the effectiveness of an evolutionary algorithm that relies completely on species selection for
evolution and on interactions among species to determine fitness. Under Resource-defined Fitness Sharing
(RFS), all individuals have the same objective fitness, but they act to reduce their shared fitness through
competition for resources. In previous studies, RFS has been used to evolve populations of mutually non-
competing (i.e., non-overlapping) shapes on shape nesting problems. In this paper we test the effectiveness
of a modified version of RFS, which we call PCSN, against three commercial software packages for shape
nesting. PCSN uses species proportions to represent a population, thereby simulating an infinitely large
population. With no discovery operators, such as mutation or recombination, evolution consists of
selection only, with all species present in the initial population. We show that on some shape nesting
problems this approach can outperform some commercial packages. In particular, PCSN nests more circles
on a fixed, polygonal substrate than do most of the commercial packages. This might be considered a
surprising result, since the algorithm is radically different from any shape nesting algorithms deployed to
date. While conventional methods place one shape at a time, the co-evolution approach attempts to place
all shapes simultaneously.
1 INTRODUCTION
Recent work (Horn, 2002, 2005) on a co-
evolutionary approach to the shape nesting problem
has yielded intriguing results on various types of
shapes. The nestings “look good”. But to date the
resource-defined fitness sharing
1
(RFS) method has
not been directly compared to other algorithms. In
this paper we document the first such experiments,
in which a modified version of RFS is directly
compared to existing algorithms in the form of
commercial software packages. The results indicate
that this new approach is competitive with all three
packages, even outperforming two of them.
Our modifications to RFS attempt to bring out
the essential co-evolutionary aspects of the
algorithm, which we believe perform subset
selection on the set of species (i.e., unique
individuals) present in the initial population, seeking
the “optimal” subset. To focus on these aspects of
co-evolution, we remove all discovery operators
1
US Patent No. 7181702.
(e.g., mutation and recombination), relying solely on
selection. We remove individuals from the
population representation, and replace them with
species proportions, implying an infinitely large
population. We call this Pure Co-evolutionary
Shape Nesting, or PCSN.
2 BACKGROUND
We discuss the shape nesting problem domain and
the application of the RFS approach to it.
2.1 The Shape Nesting Problem
The general problem at hand involves “nesting”
(that is, placing) shaped pieces on a finite substrate
so as to maximize the number of such pieces on the
substrate. The objective is often stated, equivalently,
as the minimization of “trim” (i.e., unused substrate)
(Dighe and Jakiela, 1996; Kendall, 2000). No
overlaps among the placed pieces are allowed, and
all such pieces must be placed so as to be completely
255
Horn J..
PURE CO-EVOLUTION FOR SHAPE NESTING.
DOI: 10.5220/0003089402550260
In Proceedings of the International Conference on Evolutionary Computation (ICEC-2010), pages 255-260
ISBN: 978-989-8425-31-7
Copyright
c
2010 SCITEPRESS (Science and Technology Publications, Lda.)

within the boundaries of the substrate. Figure 1
depicts a nesting of polygons within a polygon.
Shape nesting problems arise in a number of
industries, such as automotive manufacturing, in
which various shaped parts must be stamped from
sheet metal substrate, and the garment industry,
where component apparel pieces must be cut from
bolts of cloth (Kendall, 2000).
Here we assume a finite, two-dimensional
problem: a flat substrate of fixed size, and flat pieces
to be nested. We assume identical shape: there is
only one shape we are nesting. We limit ourselves to
arbitrary, (possibly non-convex) polygons for the
substrate and to identical circles for the shaped
pieces to be nested. These limitations help to reduce
the number of parameters to vary in each algorithm
when we run comparison tests, which is the main
focus of this paper.
2.2 Previous Work
Horn (2002) applied RFS to one and two-
dimensional shape nesting problems but limited his
tests to axis-aligned squares for the shaped pieces.
Horn (2005) extended the 2002 results by applying
RFS to arbitrarily shaped polygons, for both the
substrate and for the pieces to be nested. Figure 1
shows one of the nesting problems, along with one
nesting found by RFS, from the 2005 paper. More
recent work on RFS is of a theoretical nature (Horn,
2008).
Figure 1: General polygon nesting (Horn, 2005).
2.3 The RFS Algorithm
The implementation of resource-defined
fitnesssharing on which we base PCSN is the same
as used by Horn (2002, 2005).
2.3.1 Encoding the Decision Variables
For the two dimensional shape nesting problem, the
placement of a piece is made by specifying the
piece’s location and its orientation (rotation).
Because we are nesting circles in the current paper,
we only have to specify location, which will be an
(x,y) coordinate in the Cartesian plane.
2.3.2 RFS Selection
Every individual of the current population is
evaluated and assigned a fitness. As with Horn
(2002, 2005), chromosomes that specify a placement
of a piece that extends beyond the boundaries of the
substrate are assigned a fitness of 0. All “feasible”
individuals (i.e., chromosomes specifying piece
placements entirely on the substrate), receive fitness
greater than 0.
For each of the feasible individuals we compute
a shared fitness (Goldberg and Richardson, 1987)
for use in a standard selection method. In RFS, the
shared fitness for each individual is a function of the
area of the individual piece, its placement on the
substrate, and the extent to which the placed piece
overlaps with other placed pieces.
To describe the selection mechanism more
specifically, we need to define terms. Figure 2
illustrates the terms for two overlapping shaped
pieces a and b. The area of the shaped piece, which
is identical for all individuals in the population, is
used as the objective fitness of each individual: f
a
= f
b
= f
i
for all shaped pieces i. The area of
overlap between pieces a and b is defined as f
ab
, so
that 0 ≤ f
ab
≤ f
a
. Overlap is symmetric: f
ab
=
f
ba
.
Figure 2: RFS terms with non-convex polygons.
Next we define what we mean by species and
how they relate to individuals. In (Horn, 2002,
2005) the population consists of individuals. An
individual is simply a member of the population.
Two individuals from the current population may
have the exact same alleles on their chromosomes,
f
a
f
b
f
ab
( = f
ba
)
ICEC 2010 - International Conference on Evolutionary Computation
256

yet they occupy different “slots” in the population
(e.g., different elements of the data structure holding
the population) and so represent distinct individuals.
A species is considered to be a set of identical
individuals (i.e., with identical placements). Thus
unique chromosomes (x,y) map one-to-one with
unique species. There is complete overlap between,
and only between, any two members of the same
species.
Now we can give the specific form of the RFS
shared fitness f
Sh,k
calculation:
species Y
Sh,X
h
ih
i
Sh,i
XYY
X
fn
f
f
f
f
f
The upper and lower equations above are
equivalent. Both have an objective fitness in the
numerator, and a niche count, calculated over the
entire current population, in the denominator. In
Equation 1a, the summation in the niche count is
taken over the population of individuals (using the
variable h). In Equation 1b, the population is divided
into species Y. Each species consists of the set of all
individuals with the same chromosomes (from the
current population). Thus the shared fitness for any
member of a species X is equal to the objective
fitness of that species divided by the niche count for
that species, which is computed as the sum over all
species of the interaction term (f
XY
) multiplied by the
number of members in that species, n
x
. For example,
the RFS shared fitness calculation for the two
species A, B in Figure 2 (corresponding to the two
individuals a and b respectively) would be
.,
AB B
B
AB A
A
fnfn
f
f
fnfn
f
f
AB
Sh,B
BA
Sh,A
(2)
3 THE PCSN ALGORITHM
In this paper we choose to implement the RFS
approach a bit differently to take advantage of what
we perceive as the true strengths of this type of co-
evolution.
The previous section describes a more traditional
implementation in which a population of individuals
undergoes selection, mutation, and recombination.
But as noted there and in (Horn, 2008), it seems that
the RFS type of co-evolutionary selection operates
on species, as defined above. Furthermore, it seems
to be able to handle very large numbers of
individuals and species, so that we might be able to
rely on a large initial population for diversity, rather
than on discovery operators such as mutation and
crossover.
Therefore in the approach taken here, which we
call pure co-evolutionary shape nesting (PCSN), we
will (1) implement the infinite population model
(usually used for theoretical studies of an
evolutionary algorithm), dealing only with species,
not individuals, and (2) use selection only, omitting
all forms of mutation and recombination.
We maintain a population of species. For each
species we keep track of the proportion of the
population claimed by the species at a particular
generation. (Note that each species corresponds to a
unique piece placement, in this case an x,y
coordinate for a circle, as describe in the previous
section.) We apply proportionate selection to obtain
the expected proportion of the population for each
species in the following generation. Thus we model
an infinite population, at least to the extent of the
precision in our computer representation of a species
proportion.
Specifically, let p
x
(t) be the proportion of species
x at time (generation) t. Then under proportionate
selection, the expected proportion of x at time t+1 is
given by:
(3)
where f
sh,x
(t) is the shared fitness of species x at time
t, as described in the previous section. For PCSN,
we use this equation to compute the next generation.
(In generation 0, we initialize the population of
species to equal proportions).
4 EXPERIMENTS
We compare PCSN with existing implementations
of “mature” shape nesting algorithms, to get some
idea if this new approach has potential for practical
application. First we discuss the problem at hand.
(1a)
(1b)
PURE CO-EVOLUTION FOR SHAPE NESTING
257

4.1 A Nesting Problem
We consider non-convex polygon P1 defined in
Table 1. (Note that depictions of P1 in this paper
assume the origin is in the upper left corner, so that
the y-axis values increase down the page, while x-
axis values increase to the right.) The shape to be
nested is a circle of radius 72.
Table 1: Polygon P1.
(x,y) coordinates for fourteen vertices
in clockwise order of visitation
(0,56), (110,0), (254,0), (356,175), (476,142), (644,98),
(770,128), (770,196), (742,309), (619,406), (500,363),
(363,421), (350,509), (309,645), (127,673), (55,533),
(56,372).
We compare three commercial software
packages with each other and with RFS-based Pure
Co-evolutionary Shape Nesting (PCSN) on P1.
4.2 Three Software Packages
We apply the demonstration/trial versions of three
commercially available software packages that
include a shape nesting tool. All three, as well as the
PCSN program, are run on the same machine, a
Lenovo R61 laptop with a dual-core 1.8 GHz
processor and 2 GB of main memory. The three
packages are listed in Table 2.
Table 2: Commercial shape nesting software used.
Package
Name
Version Company Web Site
ArtCAM
Insignia
3.000k Delcam Plc www.artcam.com/
ProNest
8.2.1.1
MTC
Software
www.mtc-
software.com
OptiNest
2.3.1
Boole &
Partners
www.boole.eu
4.3 PCSN Setup
All runs of the PCSN algorithm use a diversity
setting of 6000 species. The species are generated at
random by drawing a minimum size bounding
rectangle around P1 and then generating integer
coordinates x and y independently and uniformly at
random within the bounding rectangle, discarding
duplicates and infeasibles (those coordinates which
would center a circle outside of P1 or intersecting a
side of P1), until 6000 unique and feasible
coordinates are found. Selection is then iterated once
per generation. Each generation we examine the set
of species whose proportion in the current
population exceeds 1/6000. When this set consists
entirely of non-overlapping circles, we stop the
algorithm and display that set of species.
4.4 Results
In this section we present separately the results of
running each of the shape nesting approaches:
PCSN and the three commercial packages, each
nesting the same shape, a circle of diameter 72 units,
in the same polygon (P1).
PSCN: Figure 3 shows that PCSN can nest 12
circles in P1.
Figure 3: PCSN fits 12 circles into P1.
Figure 4: ArtCAM nests 11 circles in P1.
ArtCAM: In Figure 4 we see the results of one run
of ArtCAM Insignia with a nesting of 11 circles.
ArtCAM’s nesting parameters require that the user
specifies which of four corners to use as the start of
nesting: upper left, upper right, lower left, and lower
ICEC 2010 - International Conference on Evolutionary Computation
258
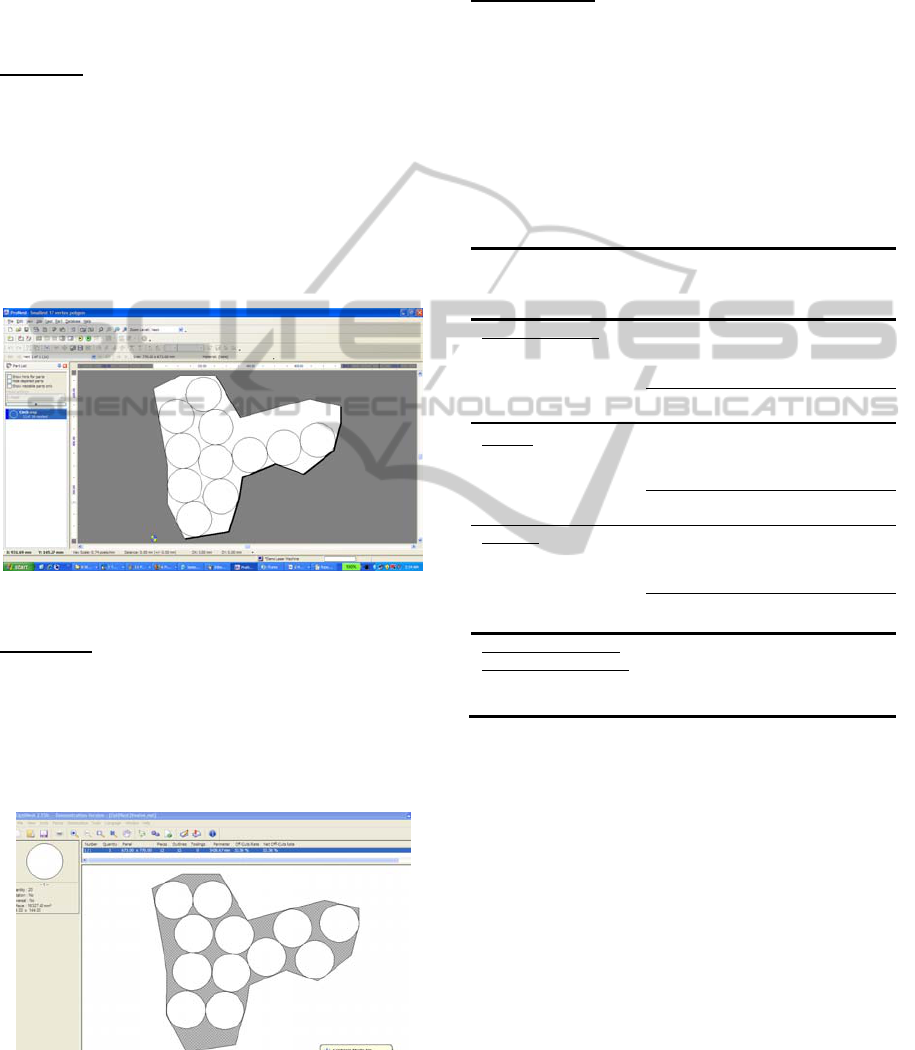
right. A second, independent nesting parameter
specifies the direction of nesting (implying a
sequential algorithm): the x-direction or the y-
direction. This yields eight possible settings of
ArtCAM’s nesting parameters. We tried all eight,
and all eight yielded an eleven circle nesting.
ProNest: Unlike ArtCAM, ProNest has many
different nesting parameters with many possible
settings. We first tried ProNest on P1 using the
initial setting of parameters as they are found when
the program is started. This resulted in a nesting of
ten circles (not shown).
We next explored the range of possible nesting
parameter settings and settled on what appeared to
be an optimal setting, resulting in the eleven circle
nesting shown in Figure 5.
Figure 5: ProNest with optimized settings fits 11.
OptiNest: With OptiNest, there are also a large
number of nesting parameters to “tune”, some of
which are continuous-valued, resulting in an infinite
number of possible parameter combinations. So we
first tried a nesting using what OptiNest names the
“default settings”. The result was an eleven circle
nesting (not shown).
Figure 6: OptiNest with optimized settings fits 12.
By spending about thirty minutes exploring the
effects of various nesting parameters (e.g.,
“subgroup size”), we settled on what appears to be
the optimal performance for OptiNest on P1. Figure
6 shows the twelve circle nesting.
SUMMARY: Table 3 summarizes these results.
The numbers in bold are the best-seen-so-far, which
is twelve circles for P1, obtained by both the PCSN
and OptiNest with optimal parameter settings. Note
that the run times are approximate, and simply show
that all four approaches are similar in execution
time. In much of the literature on shape nesting
applications, a run time of several minutes or less is
considered acceptable.
Table 3: Results on Polygon P1.
Method or Package No. of circles
nested in P1
Approximate
Run Time
(seconds)
ArtCAM Insignia
(with initial settings)
(best of all runs)
11
2
11 60
ProNest
(with initial settings)
(best of all settings)
10
1
11 5
OptiNest
(with default settings)
(with optimized settings)
11 2
12 140
Pure Co-evolutionary
Shape Nesting (PCSN)
(with diversity 10000)
12
70
5 DISCUSSION
The modified version of RFS that we introduce as
PCSN (Pure Co-evolutionary Shape Nesting)
represents a population distribution as a vector of
species proportions, thus allowing the use of a
simple replicator equation to implement evolution.
The PCSN approach has a number of possible
advantages and disadvantages but this study is
meant only to introduce the approach.
The above results on polygon P1 provide
evidence that the co-evolutionary approach to shape
nesting can sometimes perform as well as, or better
than, existing, deployed algorithms (as sampled in
the current commercial software tested here). But
the application of the commercial software packages
PURE CO-EVOLUTION FOR SHAPE NESTING
259

was not performed by someone trained in the use of
such packages. For ProNest and OptiNest in
particular, the number of parameters to tune is quite
a high. A more rigorous study is in order, in which
parameter settings are explored in a systematic
manner. (On the other hand, the fact that commercial
packages require such parameter tuning, while
PCSN does not, can be considered a strength of
PCSN in particular, and of co-evolutionary
approaches in general.)
And yet PCSN does have one critical parameter:
its diversity setting. This corresponds to the “species
population size”, that is, the number of species
represented. How high should this be set for a
particular problem? Without discovery operators
such as mutation and crossover, even if PCSN can
scale to larger (i.e., more species) populations, it is
not clear that enough diversity can be generated this
way.
And larger, more challenging problems are out
there. Nesting circles, even inside of arbitrary
polygons, is not as impressive as nesting more
complex shapes, especially shapes that are not
rotationally symmetric. Also, the numbers of circles
nested in P1 are relatively small. Typical problems
in industry involve nesting dozens or even hundreds
of shapes on one substrate. Still, the fact that PCSN
is able to select subsets of a dozen or so shapes from
a set of thousands (e.g., 6000) indicates some ability
to scale with problem size.
Clearly the use of selection alone, relying solely
on the initial population for genetic diversity, is a
radical step for a practical algorithm, but we hope
that it points out an important challenge and strength
of all cooperative co-evolution algorithms: the need
to perform subset selection from a large set. Even
with all the components of a good solution present
in the initial population, it is not an easy task to
select those components. Their quality is based
solely on their relationships with each other.
We hope also that we have shown that the
infinite population model, traditionally used in
theoretical models of evolutionary algorithms, can
be used in the algorithms themselves, in practical
implementations. Such a representation of an
evolving population could prove to be extremely
efficient, in terms of computational space and/or
time requirements, for co-evolutionary algorithms.
A final note on future work: there are many more
commercial packages. In addition, there are many
published, academic algorithms for shape nesting,
which have the advantage (over commercial
implementations) of being fully specified in
publications. This comparison study is just the
beginning.
REFERENCES
Dighe, R., Jakiela, M. J., 1996. Solving Pattern Nesting
Problems with Genetic Algorithms: Employing Task
Decomposition and Contact Detection Between
Adjacent Pieces. Evolutionary Computation, 3, 239-
266.
Goldberg, D. E., Richardson, J., 1987. Genetic algorithms
with sharing for multi-modal function optimization. In
Grefenstette, J. (Ed.), Proceedings of the Second
International Conference on Genetic Algorithms pages
(pp. 41-49). Hillsdale, NJ: L. Erlbaum Associates.
Horn, J., 2002. Resource-based fitness sharing. In
Guervós, J. J. M., Adamidis, P., Beyer, H.-G., Martín,
J. L. F.-V., and Schwefel, H.-P. (Ed.s), Parallel
Problem Solving From Nature (PPSN VII, Lecture
Notes in Computer Science, Vol. 2439, pp. 381-390).
Berlin/Heidelberg: Springer.
Horn, J., 2005. Coevolving species for shape nesting. In
Schaeffer, J. D. (Ed.), The 2005 IEEE Congress on
Evolutionary Computation (IEEE CEC 2005, pp.
1800-1807). Piscataway, NJ: IEEE Press.
Horn, J., 2008. Optimal nesting of species for exact cover
of resources: many against many. In Rudolph, G.,
Jansen, T., Lucas, S., Poloni, C. and Beume, N. (Ed.s),
Parallel Problem Solving From Nature X (PPSN X,
Lecture Notes in Computer, Vol. 5199, pp. 438-448).
Berlin/Heidelberg: Springer.
Kendall, G., 2000. Applying Meta-Heuristic Algorithms to
the Nesting Problem Utilising the No Fit Polygon.
Ph.D. thesis, University of Nottingham.
ICEC 2010 - International Conference on Evolutionary Computation
260
