
Evolution of Cooperation in N-player Social Dilemmas:
The Importance of being Mobile
Maud D. Gibbons, Colm O’Riordan and Josephine Griffith
Discipline of Information Technology, National University of Ireland Galway, Galway, Ireland
Keywords:
N-Player Social Dilemmas, Evolution of Cooperation, Contingent Mobility.
Abstract:
This paper addresses issues regarding the emergence of cooperation in evolutionary, spatial game-theoretic
simulations. In the model considered, agents participate in a social dilemma with their neighbours and have
the ability to move in response to environmental stimuli. Both the movement strategies and the game strategies
(whether to cooperate or not) are evolved. In particular, we present results that compare the outcomes using the
classical two player prisoner’s dilemma and a generalised N-player prisoner’s dilemma. We also explore the
effect that agent density (the number of agents present per cell in the world) has on the evolution of cooperation
in the environment. Finally, we discuss the movement strategies that are evolved for both cooperative and non-
cooperative strategies.
1 INTRODUCTION
Questions relating to cooperation and its emergence
have been studied in a range of domains including
economics, psychology, theoretical biology, and com-
puter science. Researchers have explored the con-
ditions necessary for cooperation to emerge among
groups or societies of self-interested agents. Social
dilemma games, such as the Prisoner’s Dilemma (Ax-
elrod, 1984), have been adopted as a succinct rep-
resentation of the conflict between individually self-
ish behaviours and collectively rational behaviours.
Evolutionary game theory has been studied since the
1980s when ideas from evolutionary theory were in-
corporated into game theory (Maynard Smith, 1982).
A variety of social dilemmas have been studied
with the majority of attention afforded to the 2-player
prisoner’s dilemma. Many variations of this game ex-
ist, which allow researchers to explore questions re-
garding cooperation in the presence of noise, trust,
spatial mechanisms and other extensions. One inter-
esting extension that has been explored in the liter-
ature is that of N-player social dilemmas (Yao and
Darwen, 1994) where N agents participate simultane-
ously in the interaction. Each agent can cooperate or
defect, and receives a reward based on the number of
cooperators present. Additionally, cooperators incur
a cost to interact while defectors do not.
In this work, we consider populations of agents
participating in both the 2-player and N-player ver-
sions of the prisoner’s dilemma. We adopt a spatial
model where agents’ interactions are defined by some
topological constraints. Much recent work has fo-
cused on the effect of such constraints (Szolnoki et al.,
2009; Ohtsuki et al., 2006; Lieberman et al., 2005).
We use a toroidal lattice where agents may interact
with their immediate eight neighbours, if any. We fur-
ther imbue the agents with the ability to move based
on environmental stimuli. The role of mobility in the
evolution of cooperation has grown in importance and
recognition in recent decades with several researchers
demonstrating its use in the promotion of cooperation
in artificial life simulations (Aktipis, 2004; Vainstein
et al., 2007). We adopt an evolutionary framework
where successive populations are evolved; the strat-
egy for interacting in the games and the mobility strat-
egy are both subject to evolution.
The N-player prisoner’s dilemma has not been
widely studied in evolutionary models where agents
are spatially situated with the inclusion of mobility.
We wish to explore if any significant differences are
prevalent between 2-player and N-player dilemmas in
this context. Furthermore, we wish to examine the
effect of varying the density of the agents in the en-
vironment, and finally we wish to analyse the move-
ment strategies evolved in these conditions.
In this paper, we show through simulation that
there is in fact a substantial difference between the
2-player and the N-player scenarios in terms of the
likelihood of cooperation emerging for varying den-
78
Gibbons, M., O’Riordan, C. and Griffith, J.
Evolution of Cooperation in N-player Social Dilemmas: The Importance of being Mobile.
DOI: 10.5220/0006052700780085
In Proceedings of the 8th International Joint Conference on Computational Intelligence (IJCCI 2016) - Volume 1: ECTA, pages 78-85
ISBN: 978-989-758-201-1
Copyright
c
2016 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved

sity levels. We demonstrate that for a range of density
levels, cooperation emerges in the N-player case.
The paper outline is as follows: the next section
discusses some related work in the field, section 3 out-
lines our model and approach, and section 4 presents
and discusses our results. Finally conclusions and
some potential future directions are presented.
2 RELATED WORK
In this section we review some of the relevantresearch
in the literature; we introduce some concepts pertain-
ing to social dilemmas and discuss some work on spa-
tial and evolutionary game theory and the role of mo-
bility.
2.1 Social Dilemma Games
Social dilemma games (most famously the prisoner’s
dilemma and its variants) have been studied in a wide
range of domains due to their usefulness in capturing
the conflict between individual and collectively ratio-
nal behaviours. The prisoner’s dilemma in the clas-
sical game is described as follows: two players make
a choice simultaneously to either cooperate or defect.
Mutual cooperation yields a reward R for both par-
ticipants. However, unilateral defection results in a
greater payoff, T, for the defector and a worse pay-
off, S, for the cooperator (the sucker’s payoff). If
both defect, both receive P as a payoff such that:
T > R > P > S.
It has been argued that the N-player variant cap-
tures a wider set of dilemmas (e.g. donating to char-
ity organisations, environmental issues etc.). In the
N-player dilemma game there are N participants, and
again, each player is confronted with a choice: to
either cooperate or defect. In one formalism of the
game (Boyd and Richerson, 1988), all players receive
a benefit based on the number of cooperators present.
Cooperators have to pay a cost. No such cost is borne
by defecting players. For instance, let B represent
some fixed benefit, N the number of players, c the
cost and i the number of cooperators. Participants re-
ceive (B × i)/N. Cooperators must pay c and thus
receive a net reward of ((B×i)/N) −c. This, or simi-
lar, formulas have been adopted in several other works
(O’Riordan and Sorensen, 2008; Yao and Darwen,
1994; Suzuki and Arita, 2003).
We represent the payoff obtained by a strategy
which defects given i cooperators as D(i) and the pay-
off obtained by a cooperativestrategy giveni coopera-
tors as C(i). Defection represents a dominant strategy,
that is, for any individual, moving from cooperation to
defection is beneficial for that player in that they still
receive a benefit without the cost:
D(i) > C(i) 0 < i ≤ N − 1
However, if all participants adopted this domi-
nant strategy, the resulting scenario would be a sub-
optimal, and from a group point of view, irrational
outcome:
C(N) > D(0)
If any player changes from defection to coopera-
tion, the society performs better:
(i+ 1)C(i+ 1) + (N− i− 1)D(i+ 1) >
(i)C(i) + (N − i)D(i)
In multi-person games, the problem of avoiding
exploitation, or free riders, is more difficult, and coop-
eration may be harder to achieve. In 2-player games,
reciprocity has been explored as a means to engender
cooperation (Nowak, 2006). However, in N-person
games reciprocity may be less advantageous. In order
for an agent to punish a defector by defecting in re-
taliation, the agent must also punish all those that did
cooperate.
2.2 Evolutionary N-player Games
There have been several other notable approaches
to exploring the N-player prisoner’s dilemma using
the tools and approaches in evolutionary game the-
ory. Yao and Darwen (Yao and Darwen, 1994) ex-
plore the effect of group size in the evolution of co-
operation. Strategies are represented using a gener-
alised form of the representation employed by Axel-
rod and Dion(Axelrod and Dion, 1988). In their ex-
periments, it is shown that cooperation can be evolved
in groups but that it becomes more difficult with in-
creasing group size.
The effects of spatial influences on the evolution
of cooperation among strategies participating in the
N-players prisoner’s dilemma is explored by Suzuki
and Arita (Suzuki and Arita, 2003). The two spatial
factors under investigation are on the scale of interac-
tion (determines which neighbours to play with) and
scale of influence (specifies which neighbouring can-
didates to choose for offspring). Results for simula-
tions involving a tit-for-tat like strategy showed that
cooperation becomes most wide-spread for a modest
value of scale of interaction and that, as the cost of
cooperation increases, the levels of cooperation de-
crease and a higher value of the scale of interaction is
found. Results also indicate that higher cooperation
levels are achieved for higher values of the scale of
influence.
Evolution of Cooperation in N-player Social Dilemmas: The Importance of being Mobile
79

2.3 Mobility
Traditional spatial models promote the evolution of
cooperation by constraining agent interactions to a
particular static topology. Previous work has inves-
tigated structures such as lattices (Nowak and May,
1992), small-world graphs (Santos et al., 2006), and
scale-free graphs (Poncela et al., 2009). However, the
inclusion of movement creates a more realistic model
by allowing agents to respond to their current neigh-
bourhood by moving within their environment.
Mobility is a form of network reciprocity (Nowak,
2006), which has gone from being perceived as a hin-
drance to the emergence of cooperation to a key con-
cept in its promotion. While unrestrained movement
can, and does, lead to the ‘free-rider’ effect (Enquist
and Leimar, 1993), allowing highly mobile defectors
to go unpunished, using simple strategy rules (Ak-
tipis, 2004; Ichinose et al., 2013) or using mobility
rates (Meloni et al., 2009; Vainstein et al., 2007) sig-
nificantly curb the free-rider phenomenon allowing
self-preserving cooperator clusters to form, and co-
operation to proliferate.
Several mechanisms for the emergence of coop-
eration exist, but all essentially express a need for co-
operators to either avoid interactions with defectors or
increase and sustain interactions with other coopera-
tors. Research in this domain is largely divided into
two categories based on authors’ definition of mobil-
ity; all movement should be random (Vainstein et al.,
2007; Meloni et al., 2009; Sicardi et al., 2009; Anto-
nioni et al., 2014), or should be purposeful or strate-
gically driven, but may indeed contain random ele-
ments (Aktipis, 2004; Helbing and Yu, 2008; Helbing
and Yu, 2009; Jiang et al., 2010; Yang et al., 2010;
Tomassini and Antonioni, 2015). Random mobility
can be used to describe the minimal conditions for
the evolution of cooperation. Alternatively, contin-
gent mobility has the capacity to be proactive. This is
where individuals deliberately seek better neighbour-
hoods, rather than simply reacting to stimuli and ran-
domly relocating.
The majority of the contingent mobility strate-
gies in the literature are hand crafted or guided by
heuristics. However, there has been some research
(Joyce et al., 2006; Gibbons and O’Riordan, 2014;
Gibbons et al., 2016) using evolutionary models to
evolve movement strategies that are conducive to the
emergence of cooperation. Ichinose et al. (Ichinose
et al., 2013) also use an evolutionary model and in-
vestigates the coevolution of migration and cooper-
ation. Agents play an N-player Prisoner’s Dilemma
game after which they move locally according to an
evolved probability vector. All agents are evolved to
collectively follow or chase cooperators. The authors
highlight the importance of flexibility in the direction
of migration for the evolution of cooperation.
Chiong et al. (Chiong and Kirley, 2012) describe a
random mobility model where a population of agents
interact in an N-player Prisoner’s Dilemma set in a
fully occupied regular lattice. Pairs of agents move
by exchanging grid positions. Mobility in this envi-
ronment is a probability function based on the time
an agent has spent in a location, and the relative fit-
ness of the agent at the destination. The agents have
a limited memory of past interactions, and past coop-
erator and defector levels. Cooperation is shown to
be promoted under a limited small set of parameters
including the cost to benefit ratio of cooperation and
the movement radius.
Most recently, Suarez et al. (Suarez et al., 2015)
present a contingent mobility model, using the N-
Player game, in which agents move toward locations
with higher potential payoff. While cooperation does
emerge, the authors do not elaborate on the specific
effects of mobility, focusing more on the impact of
the neighbourhood size.
3 METHODOLOGY
3.1 Environment & Agent
Representation
The population of agents A inhabits a toroidal shaped
diluted lattice with L × L cells, each of which can
be occupied by up to one agent. The interaction
and movement radii of agents is determined using the
Moore neighbourhood of radius one. This comprises
the eight cells surrounding an individual in a cell on
the lattice. The agents can only perceiveand play with
those within this limited radius.
Each agent is represented by a genotype, which
determines their strategy to interact with other agents
and to move in the environment. The first section of
the gene describes their strategy for playing the game:
that is to cooperate or defect and the remaining sec-
tions determine how an agent will move. The remain-
der of the genotype encodes actions for a range of
scenarios that may arise within the environment, in-
cluding: encountering a cooperator, encountering a
defector, or encountering both at once. If an agent
meets a cooperator, they have a set of potential ac-
tions. These actions are as follows: remain where
they are, move randomly,follow the cooperator or flee
from it. Similarly these potential actions are mirrored
when an agent meets a defector. The final section is
ECTA 2016 - 8th International Conference on Evolutionary Computation Theory and Applications
80

used to determine actions when an agent meets both a
defector and a cooperator. The actions are: flee from
both cooperator and defector; follow both cooperator
and defector; follow the cooperator and flee from the
defector and the converse action (flee from the coop-
erator and follow the defector). During a simulation
run, each potential action of an agent is determined
by its genotype.
At each time step, agents participate in a single
round of the Prisoner’s Dilemma with each of their
neighbours, if any. The strategy with which agents
play is fixed; either always cooperate or always de-
fect. We choose to implement pure strategies in or-
der to reduce the strategy space allowing us to more
clearly examine the effect of mobility in these experi-
ments. Agents are aware of the actions taken by their
neighbours in a single round, but these memories do
not persist. Following this interaction phase, agents
have the opportunity to take one step into an adjacent
free cell according to their movement strategy. Move-
ment will not occur if there is no adjacent free space,
or if their strategy dictates that they remain in their
current location. Isolated agents will take one step in
a random direction.
3.2 Evolutionary Dynamics
The movement strategies adopted by the population
are explored by using an ALife inspired evolution-
ary model. In a single generation, agents accumu-
late their payoffs received from playing the Prisoner’s
Dilemma with their neighbours. This is used as a
measure of fitness, and at the end of each generation,
the agents are ranked according to this score. The
bottom 20% are replaced with copies of the top 20%.
This replacement strategy was chosen as it has been
previously shown to produce a fair sampling of the
population’s fitness while still allowing for conver-
gence in a reasonable time frame. No other genetic
operators are utilized. These offspring are randomly
placed on the grid, and the other agents remain in the
same place, thus maintaining any spatial clustering
between generations. Following reproduction, the fit-
ness score of the whole population is reset and a new
generation begins.
3.3 Interaction Model
In keeping with previous work, we adopt a
well known formalism for the N-player prisoner’s
dilemma. Letting B be a constant representing social
benefit, c be the cost of cooperation and i the number
of cooperators from a group of N agents, the follow-
ing payoffs are used:
C(i) =
B× i
N
− c
D(i) =
B× i
N
The following constraints hold: B > c and both B
and c are positive values.
Considering the N-player dilemma, when N=2
and attempting to align with the classical interpreta-
tion of the 2-player prisoner’s dilemma, we also re-
quire that B < 2c. Values chosen in this research that
are in keeping with previous studies in the field are:
B = 5,c = 3.
For example, in mapping this back to the two
player games, we use the following payoff matrix:
Table 1: Prisoner’s Dilemma game matrix.
C D
C 2,2 −
1
2
,2
1
2
D
2
1
2
,−
1
2
0,0
In our simulations, we contrast scenarios with 2-
player interactions and N-player interactions. In the
N-player case, an agent particpates in the dilemma
with all of its immediate neighbours; the number of
such neighbours determines the number of partici-
pants. In the 2-player case, an agent participates in
individual 2-player games with each of its immediate
neighbours.
4 SIMULATION RESULTS
4.1 Experimental Setup
In these experiments, we run two sets of similar simu-
lations, one with 2-Player interactions the other with
N-Player interactions, comparing the respective out-
comes. It is generally accepted that when compar-
ing the two interaction models inducing cooperation
in the N-Player games is considerably harder.
The population of A = 100 agents is placed ran-
domly on the L × L torus with L = 30, the strate-
gies (whether to cooperate or to defect) are assigned
in equal proportion, and the movement strategies are
assigned randomly. A single simulation lasts 1,250
time-steps, in which the agents take 25 steps in each
of 50 generations. The distribution of spatial strate-
gies, level of cooperation, time taken for the simula-
tion to convergeon cooperation (or defection), and the
total number of interactions will all be recorded. Each
simulation will be run over a 1000 times to generate
statistically valid results.
Evolution of Cooperation in N-player Social Dilemmas: The Importance of being Mobile
81
4.2 2-Player vs. N-Player
4.2.1 Cooperative Outcomes
On average in these environmental settings, the 2-
Player interaction model is more effective at induc-
ing the spread of cooperation in a larger percent-
age of simulations. We see in Table 2 that in
roughly one third of evolutionary simulations using
2-player interaction, cooperation emerges as the out-
come, whereas when agents participate in an N-player
interaction, cooperation emerges in roughly one quar-
ter of the simulations. On average, simulations using
the 2-Player interaction model tend to converge more
quickly, and with less variance. The simulations re-
sulting in the emergence of defectors exhibit a faster
convergence and less variability in convergence speed
regardless of the interaction model.
Table 2: 2-Player vs. N-Player : % Cooperator Wins.
Avg Std Dev
2-Player 33.2% 4.2%
N-Player
25.8% 4.7%
These results are in keeping with the general con-
sensus that evolving cooperation in the N-player pris-
oner’s dilemma can be more difficult. This previous
research did not allow movement of agents, but still
captured the difficulty with N-player dilemmas where
an agent can exploit multiple participants and achieve
a considerable gain in payoff per interaction.
4.2.2 Evolved Movement Strategies
Tables 3, 4 and 5 show the movement behaviours that
are evolved for 2-player and N-player situations re-
spectively in those runs when cooperation emerges.
One hundred simulation runs resulting in cooperative
outcomes are considered.
Upon seeing a cooperator in their neighbourhood,
agents evolve to either stay where they are or to fol-
low the cooperator; this occurs in both 2-player and
N-player scenarios. When a defector is encountered,
agents have evolved to flee or adopt a random move-
ment in 75% of cases in the 2-player game and 97% of
cases in the N-player game. For the scenarios where
agents see both cooperators and defectors we see sim-
ilar behaviours being evolved. Movement behaviours
Table 3: On seeing Cooperator : % Genes Evolved.
2-Player N-Player
Random 0% 0%
Follow
15% 27%
Flee 0% 0%
Stay
85% 73%
Table 4: On seeing Defector : % Genes Evolved.
2-Player N-Player
Random 34% 22%
Follow 16% 2%
Flee
41% 75%
Stay 9% 1%
Table 5: On seeing Cooperator & Defector : % Genes
Evolved.
2-Player N-Player
FollowCFollowD 27% 3%
FollowCFleeD 44% 52%
FleeCFollowD
9% 11%
FleeCFleeD 20% 34%
that promote cooperation and avoid exploitation are
selected. We can see that cooperators who interact
using the N-Player interaction model have a greater
evolutionary incentive to be adverse to defectors.
In all cases agents learn movementbehaviours that
allow them to continue cooperative interactions and,
to a lesser extent, to avoid interactions with defectors.
Behaviours that continue defector interactions die off,
although at a slower rate. Following cooperators is
selected more quickly than fleeing from defectors.
It is important to note that the selective pressure
to avoid defectors is removed when the defectors are
replaced in the population with cooperators and hence
we do not see convergence to 100% for the genes that
promote avoiding defector interactions. Adopting a
random movement can also often have the same effect
as fleeing from or indeed following an individual.
The population did not always evolve a single
strategy; random fluctuation and lack of relevant stim-
uli resulted in simulations in which agents converged
on several strategies that were genotypically different,
but phenotypically similar.
In non-cooperative runs, defectors learned to (1)
follow cooperators, (2) flee from defectors, and to (3)
follow both cooperators and defectors.
4.3 Variation in Density
In the previous experiments, the percentage of co-
operative outcomes and the evolution of movement
strategies was a function of the agent interactions.
The ratio of cooperative interactions to other types of
interactions influences the evolutionary trajectories.
In this experiment we aim to investigate the im-
pact of the density of agents in the environment. We
define the density as D = A/L
2
where A is the size of
the population, and L is the length of the lattice grid.
Density is a function of the population size and the
size of the grid. We keep the population size constant
ECTA 2016 - 8th International Conference on Evolutionary Computation Theory and Applications
82
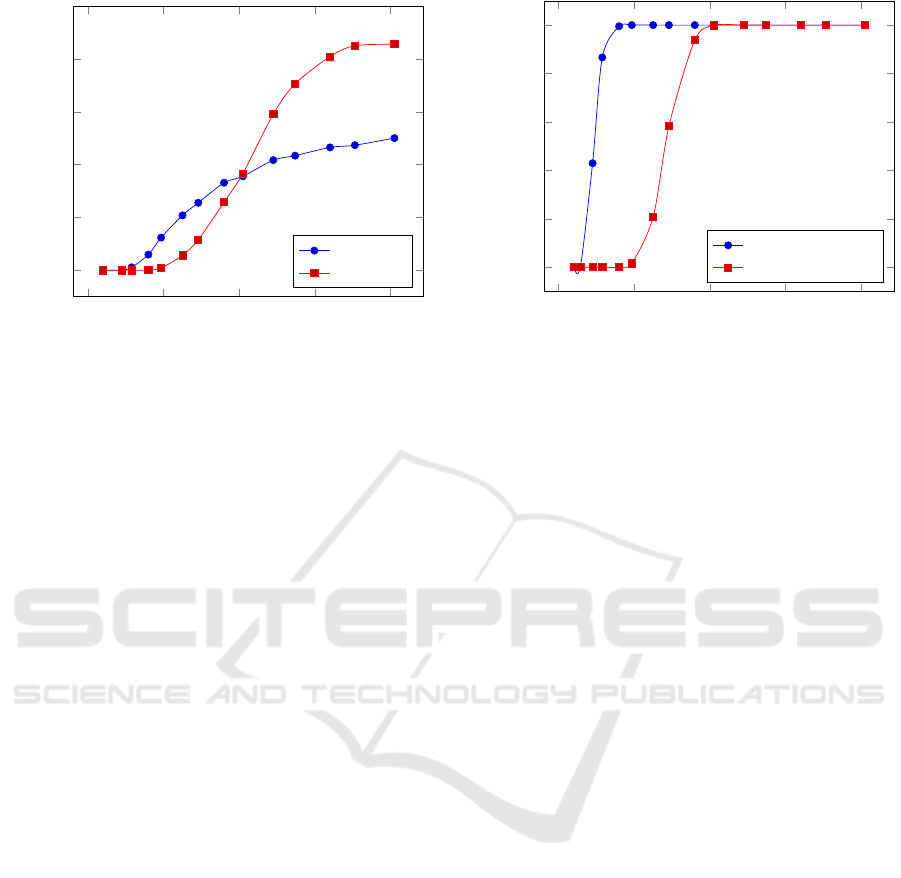
0 500 1,000 1,500 2,000
0
20
40
60
80
100
Grid Size (L
2
)
Cooperator Win %
2-Player
N-Player
Figure 1: 2-Player vs. N-Player: The percentage of simula-
tions resulting in cooperative victories as we vary the grid
density starting from random initialization.
and vary the size of the grid as a means to vary the
density.
The movement strategies of agents are randomly
initialized, the strategies for game interactions are as-
signed in equal proportions and both the movement
and interaction strategies are subject to evolution. In
one set of simulations, the population interacts using
the 2-player interaction model, and the other uses the
N-player model.
As shown in Figure 1, at the highest density level,
there is not enough space within the grid for agents
to move freely and so defection dominates in the vast
majority of simulations. These conditions echo the
traditional spatial models with an agent located in ev-
ery cell where no movement is possible. These find-
ings mirror those results with defection spreading and
dominating the population.
As the density is reduced, we see that the evo-
lutionary runs using the 2-Player interaction model
are more readily able to induce higher levels of co-
operation. However, using the 2-Player interaction
model, random initialization in low densities can only
achieve cooperation in just above 50% of simula-
tions. With these same settings the N-Player inter-
action model can induce cooperation in a far greater
percentage (80%) of runs.
For a grid size of 32x32 (1024 cells), the N-player
interactions overtakethe 2-player interaction model in
their ability to induce cooperation. This result demon-
strates that despite the difficulty of inducing coopera-
tion, cooperation emerges in N-player games, the ad-
dition of movement capabilities can support the emer-
gence of cooperation in these conditions.
0 500 1,000 1,500 2,000
0
20
40
60
80
100
Grid Size (L
2
)
Cooperator Win %
2-Player Seeded
N-Player Seeded
Figure 2: 2-Player vs. N-Player: The percentage of cooper-
ative victories, as we vary the grid density, seeding the most
prevalent evolved strategies for cooperators and defectors.
4.4 Seeding the Evolved Strategies
In our final experiment, the evolved movement strate-
gies for both cooperators and defectors are seeded in
the population and we repeat the density experiment.
In the previous experiment both movement strategies
were randomly assigned and it took several genera-
tions for movement strategies to emerge. A number of
these strategies. were identified as being favorable to
the emergence of cooperation. The aim of this exper-
iment is to explore the effect of these good strategies
when they are present in the first generation. If these
strategies help cooperators to follow each other and
form cooperative clusters, then higher levels of coop-
eration are expected across the various density levels.
Results show in both sets of simulations that the
evolved cooperator movement strategies are able to
induce cooperation for a much wider range of densi-
ties, as illustrated in Figure 2. There is a far greater
level of cooperation than that which was achieved
by either interaction model in the experiment with
random initialization. For the N-player interaction
model, once the grid size reaches 1024 (density
roughly equal to 10%), cooperation is achieved 100%
of the time. For the 2-player interaction model, this
level of cooperation is also maintained for higher den-
sity levels. The agents using the N-player model are
more hindered by the exploitative nature of defectors,
who are also using a previously evolved movement
strategy.
Evolution of Cooperation in N-player Social Dilemmas: The Importance of being Mobile
83

5 DISCUSSION
Traditionally, it has been difficult to induce cooper-
ation using the N-player Prisoner’s Dilemma. How-
ever, in our model we observe high levels of cooper-
ation in a range of settings. The incorporation of a
contingent mobility allows cooperators to cluster to-
gether, and avoid repeated defector interactions. In
forming these clusters, these agents can increase their
number of mutually cooperative interactions, thereby
boosting their score. However, these cooperative clus-
ters can be exploited by defectors unless they employ
strategies that can avoid repeated exploitativeencoun-
ters. We observe high levels of cooperation coupled
with evolved movement strategies that encourage the
formation of these larger self-preserving clusters free
from the influence of defectors.
As expected, the 2-Player interaction model was
more successful at inducing cooperation in the higher
grid densities when we evolved from random strate-
gies. This is due to the fact that while the chances
of encountering a defector are higher, they have less
of an exploitative impact on individuals or clusters
of cooperators. Surprisingly, the N-player interaction
model was significantly more successful at inducing
cooperation when the grid density was very low. We
attribute this success to the reduced chances of en-
countering a defector, and increased gains made by
mutually cooperative interactions in clusters. Addi-
tionally, single defectors benefit by being in the neigh-
bourhood of cooperators but this benefit is reduced in
the presence of other defectors.
6 CONCLUSION
In this work, we introduced a model wherein agents
inhabit a toroidal world, interact in social dilemmas,
and have the ability to move. We explored a num-
ber of scenarios involving both the 2-player and N-
player Prisoner’s Dilemma. The density of agents in
the grid was varied in a systematic manner, and its ef-
fect on the emergence of cooperation was discussed.
We present results for 2-player and N-player interac-
tion models and discuss the outcomes both in terms of
cooperation levels obtained and in terms of the move-
ment strategies selected.
Through experimentation, we show that the pres-
ence of contingent mobility strategies helps induce
cooperation in environments where the interaction
model uses the N-player Prisoner’s Dilemma. We
show that this cooperation can emerge for a range of
density levels. Furthermore, we show that seeding
a population with the previously evolved movement
strategies results in very high levels of cooperation in
both the 2-player and the N-player interaction models
for a large range of densities. These simple mobile
strategies are extremely adept at spreading coopera-
tion throughout a mobile population without the need
for complex computation, or costly memories.
Future work will involve a more thorough inves-
tigation of the nature of the cooperative clusters that
form throughout the evolutionary runs. We also wish
to explore a larger set of N-player social dilemmas
and explore a more expressive spatial topologies.
REFERENCES
Aktipis, C. A. (2004). Know when to walk away: Con-
tingent movement and the evolution of cooperation.
Journal of Theoretical Biology, 231(2):249–260.
Antonioni, A., Tomassini, M., and Buesser, P. (2014). Ran-
dom diffusion and cooperation in continuous two-
dimensional space. Journal of theoretical biology,
344:40–48.
Axelrod, R. and Dion, D. (1988). The further evolution of
cooperation. Science, 242:1385–1390.
Axelrod, R. M. (1984). The Evolution of Cooperation. Ba-
sic Books.
Boyd, R. and Richerson, P. J. (1988). The evolution of reci-
procity in sizable groups. Journal of theoretical Biol-
ogy, 132(3):337–356.
Chiong, R. and Kirley, M. (2012). Random mobility and
the evolution of cooperation in spatial N-player iter-
ated Prisoners Dilemma games. Physica A: Statistical
Mechanics and its Applications, 391(15):3915–3923.
Enquist, M. and Leimar, O. (1993). The evolution of co-
operation in mobile organisms. Animal Behaviour,
45(4):747–757.
Gibbons, M. and O’Riordan, C. (2014). Evolution of co-
ordinated behaviour in artificial life simulations. In
Proceedings of the International Conference on The-
ory and Practice in Modern Computing.
Gibbons, M. D., O’Riordan, C., and Griffith, J. (2016).
Follow flee: A contingent mobility strategy for the
spatial prisoners dilemma. In International Confer-
ence on Simulation of Adaptive Behavior, pages 34–
45. Springer.
Helbing, D. and Yu, W. (2008). Migration as a mechanism
to promote cooperation. Advances in Complex Sys-
tems, 11(4):641–652.
Helbing, D. and Yu, W. (2009). The outbreak of cooperation
among success-driven individuals under noisy condi-
tions. Proceedings of the National Academy of Sci-
ences of the United States of America, 106(10):3680–
3685.
Ichinose, G., Saito, M., and Suzuki, S. (2013). Collective
Chasing Behavior between Cooperators and Defec-
tors in the Spatial Prisoner’s Dilemma. PLoS ONE,
8(7):28–31.
ECTA 2016 - 8th International Conference on Evolutionary Computation Theory and Applications
84

Jiang, L.-L., Wang, W.-X., Lai, Y.-C., and Wang, B.-H.
(2010). Role of adaptive migration in promoting
cooperation in spatial games. Physical Review E,
81(3):036108.
Joyce, D., Kennison, J., Densmore, O., Guerin, S., Barr, S.,
Charles, E., and Thompson, N. S. (2006). My way
or the highway: a more naturalistic model of altruism
tested in an iterative prisoners’ dilemma. Journal of
Artificial Societies and Social Simulation, 9(2):4.
Lieberman, E., Hauert, C., and Nowak, M. A. (2005).
Evolutionary dynamics on graphs. Nature,
433(7023):312–316.
Maynard Smith, J. (1982). Evolution and the theory of
games. Cambridge University Press, Cambridge; New
York.
Meloni, S., Buscarino, A., Fortuna, L., Frasca, M., G´omez-
Garde˜nes, J., Latora, V., and Moreno, Y. (2009). Ef-
fects of mobility in a population of prisoner’s dilemma
players. Physical Review E - Statistical, Nonlinear,
and Soft Matter Physics, 79(6):3–6.
Nowak, M. A. (2006). Five rules for the evolution of coop-
eration. Science, 314(5805):1560–3.
Nowak, M. A. and May, R. M. (1992). Evolutionary games
and spatial chaos. Nature, 359(6398):826–829.
Ohtsuki, H., Hauert, C., Lieberman, E., and Nowak, M. A.
(2006). A simple rule for the evolution of co-
operation on graphs and social networks. Nature,
441(7092):502–505.
O’Riordan, C. and Sorensen, H. (2008). Stable coopera-
tion in the N-player prisoners dilemma: The impor-
tance of community structure. In Adaptive Agents and
Multi-Agent Systems III. Adaptation and Multi-Agent
Learning, pages 157–168. Springer.
Poncela, J., G´omez-Garde˜nes, J., Flor´ıa, L. M., Moreno,
Y., and S´anchez, A. (2009). Cooperative scale-free
networks despite the presence of defector hubs. EPL
(Europhysics Letters), 88(3):38003.
Santos, F., Rodrigues, J., and Pacheco, J. (2006). Graph
topology plays a determinant role in the evolution of
cooperation. Proceedings of the Royal Society B: Bi-
ological Sciences, 273(1582):51–55.
Sicardi, E. A., Fort, H., Vainstein, M. H., and Arenzon, J. J.
(2009). Random mobility and spatial structure often
enhance cooperation. Journal of theoretical biology,
256(2):240–246.
Suarez, D., Suthaharan, P., Rowell, J., and Rychtar, J.
(2015). Evolution of cooperation in mobile popula-
tions. Spora-A Journal of Biomathematics, 1(1):2–7.
Suzuki, R. and Arita, T. (2003). Evolutionary analy-
sis on spatial locality in n-person iterated prisoner’s
dilemma. International Journal of Computational In-
telligence and Applications, 3(02):177–188.
Szolnoki, A., Perc, M., Szab´o, G., and Stark, H.-U. (2009).
Impact of aging on the evolution of cooperation in the
spatial prisoners dilemma game. Physical Review E,
80(2):021901.
Tomassini, M. and Antonioni, A. (2015). L´evy flights and
cooperation among mobile individuals. Journal of the-
oretical biology, 364:154–161.
Vainstein, M. H., Silva, A. T. C., and Arenzon, J. J. (2007).
Does mobility decrease cooperation? Journal of The-
oretical Biology, 244(4):722–728.
Yang, H.-X., Wu, Z.-X., and Wang, B.-H. (2010). Role of
aspiration-induced migration in cooperation. Physical
Review E, 81(6):065101.
Yao, X. and Darwen, P. J. (1994). An experimental study
of N-person iterated prisoners dilemma games. Infor-
matica, 18(4):435–450.
Evolution of Cooperation in N-player Social Dilemmas: The Importance of being Mobile
85
