
Comparing Small Population Genetic Algorithms over Changing
Landscapes
Michael Curley and Seamus Hill
College of Engineering and Informatics, National University of Ireland Galway, Ireland
Keywords:
Genetic Algorithms, Changing Landscapes, Population Size, Adaptability.
Abstract:
This paper examines the performance and adaptability of a number of small population Genetic Algorithms
(GAs) over a selection of dynamic landscapes. Much of the research in this area tends to focus on GA
with relatively large populations for problem optimisation. However there is research, which suggests that
GAs with smaller populations can also be effective over changing landscapes. This research compares the
performance and adaptability of a number of these small population GA over changing landscapes. With small
population GAs, convergence can occur quickly, which in turn affects the adaptability of a GA over dynamic
landscapes. In this paper five GA variants using small population sizes are run over well-known unimodal and
multimodal problems, which were tailored to produce dynamic landscapes. Adaptability within the population
is considered a desirable feature for a GA to optimise a changing landscape and different methods are used
to maintain a level of diversity within a population to avoid the problem of premature convergence, thereby
allowing the GA population adapt to the dynamic nature of the search space. Initial results indicate that small
population GAs can perform well in searching changing landscapes, with GAs which possess the ability to
maintain diversity within the population, outperforming those that do not.
1 INTRODUCTION
Genetic Algorithms (GAs) (Holland, 1992) (Gold-
berg, 1989) are search algorithms based on Darwinian
principals of survival of the fittest. GAs are popula-
tion based optimisers and as population evolve from
generation to generation they tend to become homo-
geneous as the population converges. GAs are widely
used for various optimisation problems, however op-
timisation over changing landscapes, can prove more
difficult compared to that of static landscapes as the
populations needs to be able to adapt and escape local
optima as the landscape changes. Therefore, if popu-
lations tend to converge on a particular solution, a GA
needs to be able to evolve its population to respond to
changes over a changing landscape. Because of this,
the ability for the GA population to adapt to changes
in the search space is critical in optimising changing
landscapes. Adaptability (Mori and Kita, 2000)(Kr-
ishnakumar, 1990)(Mumford, 2004) is considered to
play an important role in how GA searches through a
changing landscapes. Associated with adaptability is
the maintenance of diversity within the population.
With this in mind, a decision needs to be taken
whether it is better to use a standard GA and restart
the population when the landscape alters or to use a
GA which maintains an element of diversity within
the population. Research such as (Cobb and Grefen-
stette, 1993; Krishnakumar, 1990; Grefenstette et al.,
1992; Goldberg and Smith, 1987) looked at station-
ary and non-stationary landscapes and found that GAs
were able to optimise functions over both landscapes,
with the maintenance of population diversity being a
critical feature. Population size is also an important
consideration in the optimisation of a changing land-
scape. Researchers such as (Ahn and Ramakrishna,
2002) (Grefenstette, 1986) (Whitely, 1989) (Krish-
nakumar, 1990) found small populations to be just
as effective in optimizing certain landscapes, thereby
avoiding the additional overhead associated with a
larger population.
This paper examines and compares the perfor-
mance and adaptability over changing landscapes, of
a number of GA implementations using small pop-
ulations. The motivation is to compare the perfor-
mance and adaptability of a number of small popu-
lation GAs over changing landscapes in order to ex-
amine the impact of diversity maintenance and deter-
mine whether restarting a standard GA or the inclu-
sion of a diversitymaintenance technique is most ben-
Curley M. and Hill S.
Comparing Small Population Genetic Algorithms over Changing Landscapes.
DOI: 10.5220/0006497802390246
In Proceedings of the 9th International Joint Conference on Computational Intelligence (IJCCI 2017), pages 239-246
ISBN: 978-989-758-274-5
Copyright
c
2017 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved

eficial over the given landscapes. The contribution is
to develop an insight into the ability of various small
population GAs to adapt and optimise changing land-
scapes by comparing their performance over specifi-
cally designed landscapes. Initial results indicate that
the use of particular diversity maintenance techniques
in small population GAs, proves beneficial over the
chosen landscapes. The paper is laid out as follows:
Section 2 examines the background literature. Sec-
tion 3 outlines the GAs and the test suite used in the
experiments. Section 4 describes the findings, Section
5 concludes and Section 6 outlines future work.
2 BACKGROUND
In relation to population diversity and convergence,
De Jong (DeJong, 1975) set a threshold value of
95% similarity between loci alleles to indicate con-
vergence. By examining fluctuations in online per-
formance we can get a indication of the level of con-
vergence within the population. Work carried out by
Mahfoud (Mahfoud, 1995) defined convergence as
occurring when the average population fitness value
for the previous 4 generations is greater than the av-
erage fitness value for the present generation. Allen
et al. (Allen and Karjalainen, 1999) used a simple ap-
proach to measure convergence and defined that if no
progress has been made after 25 generations then the
GA has converged. The approach used is this research
is Allen’s method because of its simplicity; further-
more this method give the GA a better opportunity of
avoiding getting caught in local optima in the land-
scapes used in this research.
GA theory would suggest that larger GA popula-
tions should maintain an element of diversity within
the population for a longer period when compared
to small population GAs (Leung et al., 1997) (Gold-
berg and Deb, 1991). If diversity can be maintained,
the additional genetic material or building blocks
should assist in the adaptability of the GA. How-
ever, this comes at a cost of time and complexity as
the larger population takes longer to search through
(Ahn and Ramakrishna, 2002) there exists a trade
off between the advantages of having a large popu-
lation in terms of searching a search space and the
associated additional overhead. Research by Gref-
fenstette (Grefenstette, 1986) and also by Whitley
(Whitely, 1989), found that smaller population sizes
with slightly higher crossover and mutation rates to be
just as affective when searching a landscape. The mu-
tation operator plays a role in introducing diversity by
allowing a population which has converged on a par-
ticular solution, to open up the landscape to further
exploration. However, there is a limit to it’s ability to
introduce diversity, before it becomes a random walk.
3 EXPERIMENTAL SET UP
In this paper we compare the performance and adapt-
ability of GAs using small populations over changing
landscapes. We contrast the results of a simple GA
(SGA) (Holland, 1992) to four GA variations: GA
with Elitism (GAE), Immigration GA (GAI), Micro
GA (MGA) and Diploid GA (DGA). The GA with
elitism (GAE) (Goldberg, 1989) retains the fittest
chromosomesfrom generation to generation. As there
is no guarantee of a chromosome surviving selec-
tion, with a standard GA, elitism guarantees that the
fittest individual will be maintained in the population.
The immigration GA (GAI), implemented as outlined
by (Yang, 2008), results in the best solution being
maintained in the population, while the worst indi-
vidual is replaced by a random immigrant. The Mi-
cro GA (MGA) (Krishnakumar and Bailey, 1990) re-
tains the best individual from generation to genera-
tion and once convergence is detected, the remainder
of the population is randomly initialised. Finally, the
Diploid GA (DGA) used in this research is as outlined
in (Goldberg and Smith, 1987).
3.1 Test Suite
De Jong(DeJong, 1975) used the Sphere function and
Shekel’s function in his paper and these functions
were used to test a number of parameters at differ-
ent values. Population size, crossover and mutation
rates were examined at different levels and the results
produced from these experiments lay the foundation
parameters which are still in use today on these func-
tions. The Sphere function is used to test the general
efficiency of a GA and Shekel’s function is used to
test the GAs ability to avoid getting caught in the 25
local optima or foxholes that are present in that land-
scape. DeJong found that the GA was able to optimise
both landscapes and the that the values of mutation
population and crossover values play an important in
how efficiently a GA optimises a landscape. Other re-
searchers such as Digalakis et al. (Digalakis and Mar-
garitis, 2002) carried out these experiments and were
able to optimise the Sphere and Shekel’s function by
using DeJong parameter values.
3.1.1 Sphere Function
According to DeJong (DeJong, 1975) the sphere func-
tion is a unimodallow-dimensionalquadratic function

with a minimum of zero, in other words it contains no
local optima. The Sphere Model (DeJong, 1975) is
relatively easy to optimise as it is continuous, con-
vex and unimodal. This function is normally used to
measure the efficiency of a particular algorithm. The
Sphere function is represented as follows:
f(x) =
d
∑
i=1
x
2
i
where: min( fx) = fx(0, .. . , 0) = 0 and d is the num-
ber of dimensions, −5.12 ≤ x
i
≤ 5.12 and for all
i = 1, . . . , d. The changing environment experiments
allow the GAs to search the landscape defined by the
Sphere model, with the function being inverted after
100 generations. By changing the fitness function in
this manner, what was the global optimum becomes
the furthest point from the new global optimum.
3.1.2 Shekel’s Function
Shekel’s function is a continuous non-convex, non
quadratic two-dimensional multimodal function with
25 local minima based at the points. This 2-
dimensional function contains 25 different foxholes,
each varying in depth, surrounded by relatively flat
surfaces. Shekel’s Foxhole function has the following
definition:
f(x) =
"
1
500
+
25
∑
j=1
1
j +
∑
2
i=1
(x
i
− a
ij
)
6
#
−1
where:
−65.536 ≤ x
i
≤ 65.536
a
ij
=
−32 −16 0 16 32 −32 .. . 0 16 32
−32 −32 −32 −32 −32 −16 . . . 32 32 32
Shekel’s Foxhole is a minimising problem, where
f(x) = f(0, . . . , 0) = f(−32, −32) ≈ 1. At generation
500 the objective function is inverted, meaning that
the old global optimum is now the furthest point from
the new global optimum.
3.2 Parameter Settings
Parameter setting in GA is a key component in get-
ting the algorithm to operate in an efficient and effec-
tive manner. Researchers have come up with slightly
different parameters over time but are problem de-
pended. The values used in this research are taken
from Grefenstette (Grefenstette, 1986) and found
that smaller population sizes with slightly higher
crossover and mutation rates to be just as positive to
converge on a solution and with quicker results time
due to less processing time.
Table 1: Parameter Settings.
Parameter settings Changing Sphere Changing Shekels
population size 30 30
crossover 0.7 0.7
mutation 0.01 0.01
generations 200 1000
runs 20 20
4 FINDINGS
This section outlines the results for each of the
GAs over the changing Sphere function and chang-
ing Shekel’s function. With graphs plotting the On-
line (average) and Offline (best) results. As these are
minimising problems the results tend to zero on the
graph as the GA evolves. The dynamic Sphere func-
tion experiments were run over 200 generations, with
the objective function altering at generation 100. The
dynamic Shekel’s function experiments ran for 1000
generations, with the fitness function changing at gen-
eration 500. A two sided paired Wilcoxon test was
carried out on the offline results for each of the exper-
iments to access whether the population means dif-
fer for the variant GA when compared to the SGA.
Results were considered to be statistically significant
with a Pvalue < 0.005.
4.1 SGA (SGA) Changing Sphere
Function Results
The SGA results indicate that the algorithm had lit-
tle difficulty in solving the problem, both before and
after the landscape change. Figure 1 illustrates both
the online and offline performance. As shown in Ta-
ble 2, the SGA located the global optimum value of
0, both before and after the change. The results also
indicate that the population converges quite quickly,
shown by the closeness of the online and offline re-
sults. However, as the population had converged on
chromosomes representing the original global opti-
mum, once the objective function altered, this turns
what were good chromosomes with high fitness val-
ues, into very poor solutions. This change in the land-
scape is designed to examine the GAs ability to adapt
and locate the new global optimum. Optimisation
takes longer at this point, as the population is made
up of solutions that are much further away from the
solution than the initialised population at the start.
The SGA adapting to the change in the objective
function and searching for the new global optimum
is shown by the slight increase in distance between
the online and offline results, as the population be-
gins to move away from the area of the search space
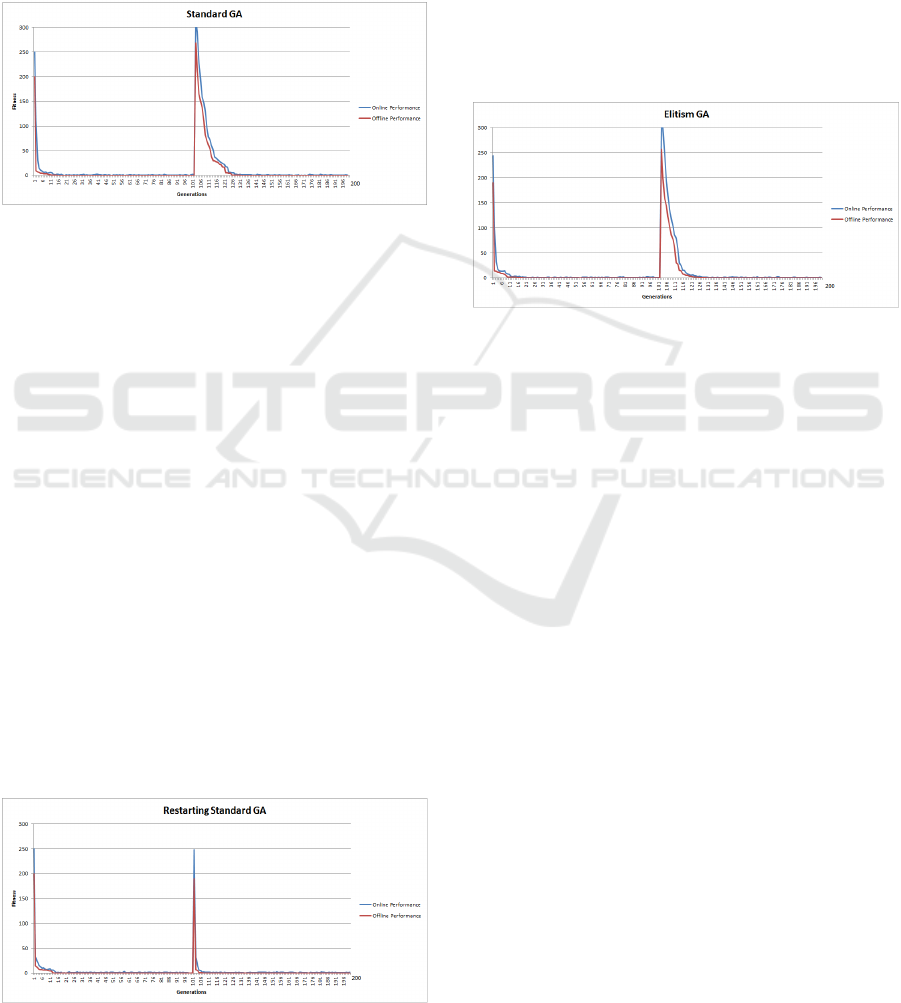
where is had converged. Following the change in
the fitness function, the SGA eventually locates the
global optimum (see Table 2) and begins to converge
as the distance between the offline and online results
decreases. Overall the SGA, using a small popula-
tion, had little difficulty in optimising the landscape
both before and after the fitness solution representa-
tions were changed, this is largely due to the nature of
the Sphere function.
Figure 1: SGA Changing Sphere Function.
4.2 SGA Restart (SGAR) Changing
Sphere Function Results
Figure 2 outlines the online and offline performance
of restarting or reinitialising the population of the
SGA when the landscape alters. The results indicate
that in the initial phase of the search, the performance
of the SGAR is similar to that of the SGA. However,
once the landscape changes, the early online and of-
fline results of the SGAR are better than those of the
SGA. This appears to indicate that the SGAR, through
random initialisation, appears find it easier to locate
the new global optimum. One reason for this is that in
the random initialisation of the population, conducted
after the change in the objective function, leaves the
population, on average, closer to the new global op-
timum, rather than converged around the old global
optimum as is the case for the SGA. Table 2 indi-
cates that in terms of locating the global optimum
the performance of the SGAR is comparable with that
of the SGA, but marginally better after the landscape
change. Comparing the offline results of the SGAR to
Figure 2: SGA Changing Sphere Function.
those of the SGA, the results were shown to be statis-
tically highly significant (p − value < 2.2e
16
).
4.3 GA with Elitism (GAE) Changing
Sphere Function Results
Figure 3 contains the offline and online results for
the GAE. Overall, the GAE’s online and offline per-
formances appear marginally better than that of the
SGA, due to the introduction of elitism (see Table 2).
The offline results of the SGA compared to GAE were
shown, using to be statistically highly significant with
a p − value of 2.817e
−08
.
Figure 3: GA with Elitism (GAE) Dynamic Sphere Func-
tion.
4.4 Immigration GA (GAI) Changing
Sphere Function Results
The GAI results (see Figure 4), the global optimum
value of 0 was located on average, at generation 4.
Once the landscape altered, the offline global opti-
mum of 0 was located at generation 138. The GAI
used in this paper, removes the worst solution in each
generation and replaces it with the best solution from
the previous generation. This allows the GAI to insert
and remove solutions based on the fitness function for
the particular landscape. The results, illustrated in Ta-
ble 2, indicate that over the changing Sphere function,
the Immigration GA outperforms the SGA, SGAR
and GAE, before and after the landscape change. Al-
though the offline results compare favourably, it is in-
teresting to note that the online results shown in Fig-
ure 4, after the landscape change, illustrate the GAI’s
ability to adapt through the introduction of diversity
into the population. This is shown in the graph by
the increase in distance between the online and offline
lines. The offline results of the SGA and GAI for the
Dynamic Sphere experiment were statistically highly
significant (p − value < 2.2e
16
).
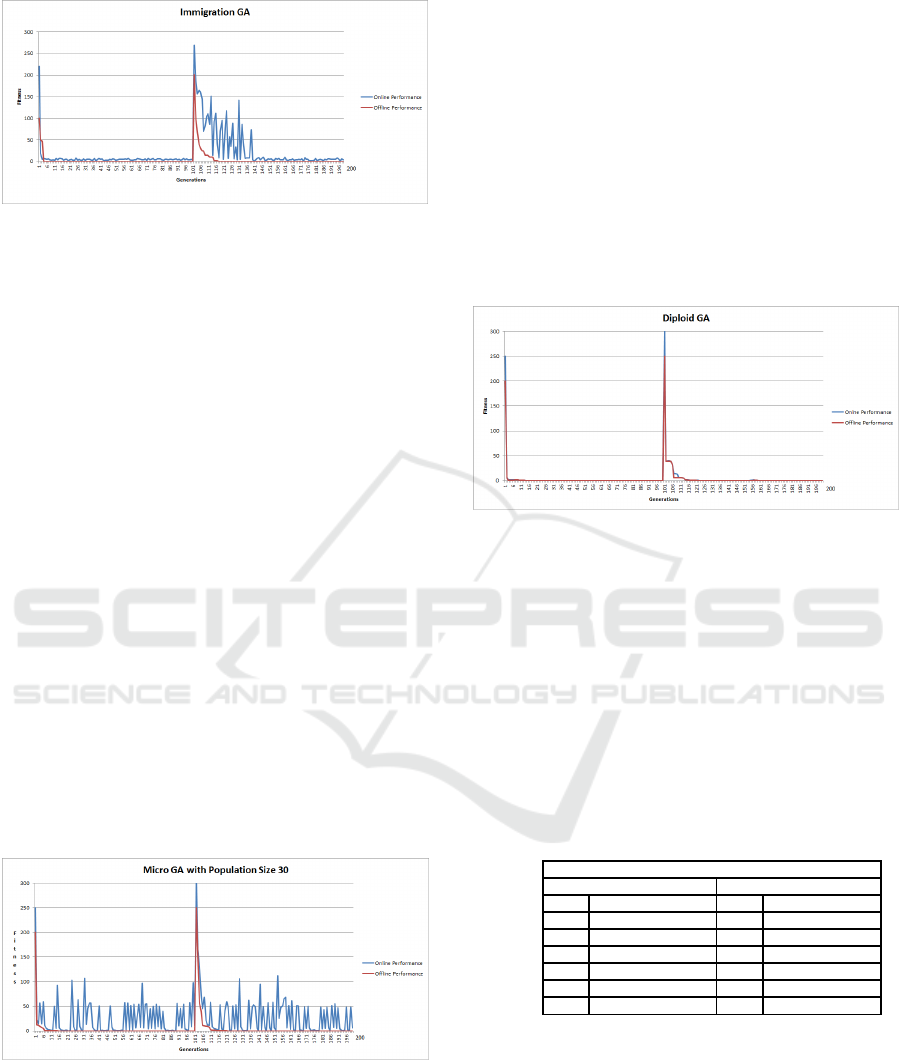
Figure 4: Immigration GA (GAI) Changing Sphere Func-
tion.
4.5 Micro GA (MGA) Changing Sphere
Function Results
The results of the MGA are shown in Figure 5. The
results of the MGA, in locating the global optimum,
indicate an improvement over the SGA, SGAR and
GAE algorithms, both before and after the objective
function changes (see Table 2). The restarting mech-
anism and elitism of the MGA appears to allowed
the algorithm adapt much quicker to the altered land-
scape, thus optimization the changing Sphere prob-
lem in the least amount of generations. The of-
fline MGA result, were statistically highly significant
when compared to the offline SGA results, with a
p − value = 0.0009585. The other interesting fea-
ture of the MGA relates to the online performance,
both before and after the landscape alters, the MGA’s
online performance suggests a significant degree of
diversity is maintained in the population. This dif-
fers dramatically from the SGA’s online performance,
which suggests a large degree of convergence. This
suggests that the ability to maintain diversity within
the population, as implemented by the MGA, assists
in adapting to the landscape change.
Figure 5: Micro GA (MGA) Changing Sphere Function.
4.6 Diploid GA (DGA) Changing
Sphere Function Results
The DGA results, seen in Figure 6, shows the offline
value of 0 was found at generation 5. At generation
100 the landscape alters, with the offline value of 0
located at generation 141. The results indicate that
over the dynamic Sphere landscape the DGA outper-
formed the SGA and the offline results, when com-
pared were statistically significant with a p−value =
0.03033. When looking at the results of the DGA,
it appears that the maintenance of diversity associ-
ated with the diploid structure, assists in adapting to a
changing landscape. There is however, an additional
overhead associated with the diploid structure com-
pared to haploid GA. Further, in terms of the fitness
of each of the diploid solutions, there is a large degree
of convergence as indicated by the online results.
Figure 6: Diploid GA (DGA) Changing Sphere Function.
Table 2 outlines the results of the changing Sphere
Function experiments, identifying whether the global
optimum was located both before and after the land-
scape changed. The results indicate that the most
adaptable small population GAs were the GAI, MGA
and DGA. However, in terms of adaptability over the
dynamic Sphere function landscape, all of the GAs
succeeded in locating the global optimum. One rea-
son for this is the relative ease of the Sphere function,
however using the Sphere function indicates the effi-
ciencies of each GA variant.
Table 2: Changing Sphere Optimisation
Changing Sphere Function Experiments
Before Landscape Change After Landscape Change
Gen. Optimum Gen. Optimum
SGA 85 Yes 170 Yes
SGAR 90 Yes 165 Yes
GAE 58 Yes 161 Yes
GAI 4 Yes 138 Yes
MGA 34 Yes 136 Yes
DGA 5 Yes 141 Yes
4.7 SGA Changing Shekel’s Function
Results
The SGA results (Figure 7) evolved for a pre-
determined number of generations and the problem
was optimised in generation 470. The results indi-
cate that Shekel’s function proved more difficult for
the SGA compared to the Sphere function. At gen-
eration 500 the objective function inverted causing
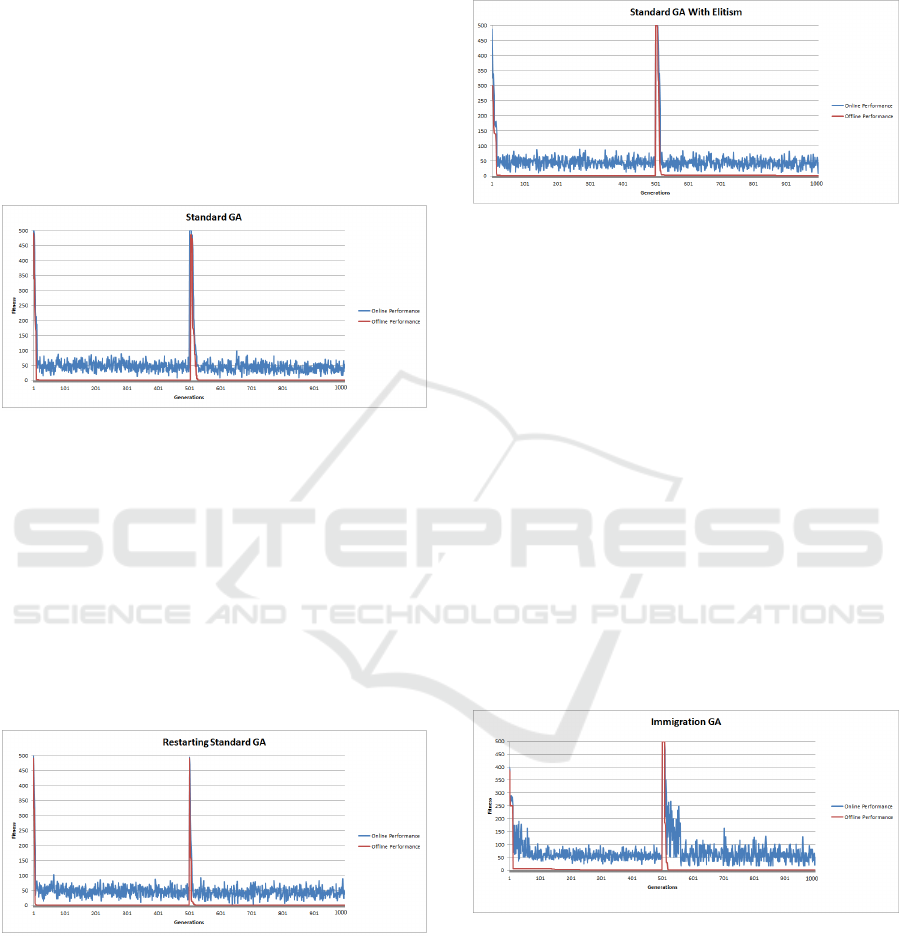
the landscape to change. At this point the population
has evolved around the old global optimum which is
much further away from the solution than the initial-
ized population, thereby making the adaptation more
difficult. The SGA however, managed to locate the
new global optimum at generation 990. The SGA lo-
cated the global optimum both before and after the
landscape change (see Table 3). The online results
are interesting as, they illustrate the increase in diffi-
culty of Shekel’s function in comparison to the Sphere
function, with slightly less convergence within the
population.
Figure 7: SGA Dynamic Shekel’s Function.
4.8 SGA Restart (SGAR) Dynamic
Shekel’s Function Results
Figure 8 illustrates the results of restarting the SGA
when the landscape alters. The global optimum was
located before and after the change in the objective
function, as shown in Table 3. The performance was
in line with that of the SGA, with a minor improve-
ment. The offline results, compared to those of the
SGA were statistically highly significant.
Figure 8: SGA Dynamic Sphere Function.
4.9 GA with Elitism (GAE) Dynamic
Shekel’s Function Results
The GAE results (see Figure 9) shows the global op-
timum being discovered before and after the land-
scape changed. The performance was largely similar
to that of the SGA and SGAR, this is shown in Table
3. The offline performance results of the GAE and
SGA were shown to be statistically highly significant
(p − value < 2.2e
−16
).
Figure 9: GA with Elitism - Dynamic Shekel’s Function.
4.10 Immigration GA (GAI) Dynamic
Shekel’s Function Results
The online and offline results of the GAI experiments
are illustrated in Figure 10. The results indicate that
there is a higher degree of diversity maintained within
the population in comparison to the SGA, SGAR and
GAE (illustrated by the online performance). The of-
fline performance showsthe global optimum being lo-
cated in both phases of the search, see Table 3. The
performance of the GAI showed an improvement over
that of the SGA beforethe objectivefunction changes.
After the change, however the results indicate that the
GAI adapted quicker in locating the new global opti-
mum. This is due to the ability of the GAI to main-
tain diversity into the population. The offline per-
formances of both algorithms was statistically highly
significant with a probability value < 2.2e
16
.
Figure 10: Immigration GA (GAI) Dynamic Shekel’s Func-
tion.
4.11 Micro GA (MGA) Dynamic
Shekel’s Function Results
Figure 11 shows the offline and online results for
MGA. The offline performance illustrates the success
in locating the global both before and after the land-
scape changes. The online performances shows the
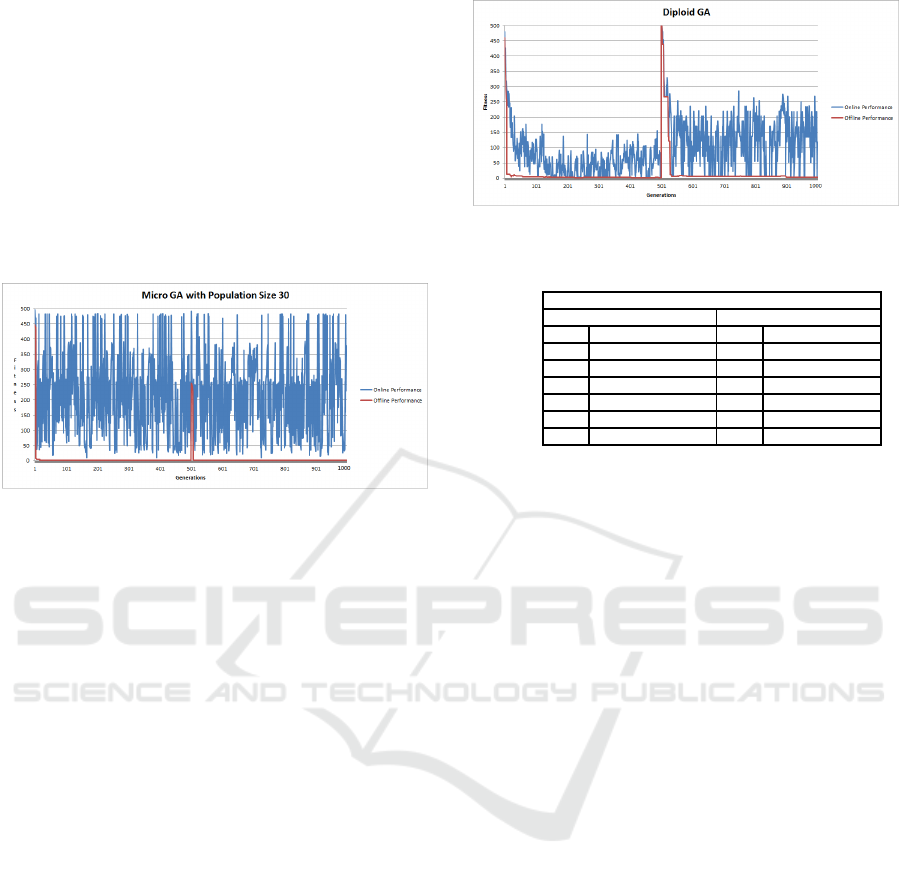
large amount of diversity being introduced into the
population as the average fitness values vary errati-
cally. However, Table 3 indicates that the MGA ex-
hibited the ability to succeed in the search, locating
the global optimum before and after the alteration, in
the fewest number of generations, compared to the
other GA variations. The ability to continuously in-
troduce new individuals into the population, appears
to beneficial over the given landscape. Statistically,
the offline performance of the MGA compared to that
of the SGA was shown to be highly significant with a
p− value < 2.2e
16
.
Figure 11: Micro GA Dynamic Shekel’s Function.
4.12 DGA Dynamic Shekel’s Function
Results
For the DGA, the offline global optimum was found
at generation 450. At generation 500, when the land-
scape was altered, the new global optimum was lo-
cated during generation 900 (see Table 3). When ex-
amining Figure 12, the offline performance is simi-
lar to that of the SGA, SGAR and GAE. The Diploid
GA maintained a high level of diversity within the
population due to the double chromosome structure.
This can be seen through the fluctuating offline per-
formance associated with the DGA, particularly after
the landscape changes as it attempts to adapt to the
changing objective function. However, in terms of
adapting to the more difficult Shekel’s function, the
DGA did not perform as well as the other diversity
maintenance GA variations, the MGA and GAI. One
reason for this may be due to the diploid structure,
which through its dominance scheme, struggles with
the landscape in question. Again, when comparing
the results of the offline performance of the SGA to
those of the DGA, the results were found to be statis-
tically highly significant with a p − value < 2.2e
16
.
A summary of the Dynamic Shekel’s function ex-
periments is shown in Table 3. The results indicate
that although all of the GAs managed to locate the
global optimum, before and after the landscape al-
tered, the GAI and MGA were the most successful.
Figure 12: Diploid GA - Dynamic Shekel’s Function.
Table 3: Dynamic Shekle’s Optimisation.
Dynamic Shekel’s Function
Before Landscape Change After Landscape Change
Gen. Optimum Gen. Optimum
SGA 470 Yes 990 Yes
SGAR 460 Yes 950 Yes
GAE 450 Yes 938 Yes
GAI 437 Yes 622 Yes
MGA 23 Yes 614 Yes
DGA 450 Yes 900 Yes
5 CONCLUSION
The results presented indicate that the small pop-
ulation GAs variations were capable of optimising
both the dynamic Sphere landscape and the dynamic
Shekel’s landscape. The research suggests, as ex-
pected, that GAs which maintained diversity into the
population, were quicker to adapt when a changing
environment was presented. However, what was in-
teresting is that each of the GAs had a relatively
small population to work with, in comparison to pre-
vious research. The findings of the DGA experiments
tended to concur with previous research where Gold-
berg (Goldberg and Smith, 1987) suggested not much
improvement over the SGA in terms of number of
generations required to optimize the landscapes. The
greater level of diversity in the population produced
disappointing results in the changing landscape given
the additional overhead required.
The dynamic Sphere results indicate that the GAs
associated with diversity maintenance,that is the GAI,
MGA and DGA all possess an ability to adapt to a
changing landscape and outperform the SGA, SGAR
and GAE. The changing Shekel’s function results
show some evidence that the GAE had more diffi-
culty in getting out ofthe local optima when compare
to the GAI and MGA. The algorithm would continu-
ously keep adding the best chromosome to the pop-
ulation and so may end up getting caught in some of
the foxholes of the changing Shekel’s landscape while
the algorithm optimized the solution it required more
generations to work through the landscape.

Overall the MGA exhibited the greatest ability to
adapt over the more difficult dynamic Shekel’s land-
scape, in comparison to the other GAs. The MGA
also proved a useful GA variation over the dynamic
Sphere landscape, producing, along with the GAI,
the lowest number of generations required in order
to adapt to the change in objective function. The re-
sults seem to imply that over changing landscapes, the
combination of small populations and diversity main-
tenance can prove successful. This may suggest that
diversity maintenance techniques coupled with small
populations, can reduce the computational overhead
and assist GAs over changing landscapes, particularly
as the level of difficulty increases.
6 FUTURE WORK
Future work includes further examination into the
coupling of small population GA and diversity main-
tenance techniques over more difficult and varied dy-
namic landscapes to see if the maintenance of diver-
sity within a small population GA continues to offer
an advantage.
REFERENCES
Ahn, C. W. and Ramakrishna, R. S. (2002). A genetic al-
gorithm for shortest path routing problem and the siz-
ing of populations. IEEE transactions on evolutionary
computation, 6(6):566–579.
Allen, F. and Karjalainen, R. (1999). Using genetic algo-
rithms to find technical trading rules. Journal of fi-
nancial Economics, 51(2):245–271.
Cobb, H. G. and Grefenstette, J. J. (1993). Genetic algo-
rithms for tracking changing environments. Technical
report, DTIC Document.
DeJong, K. A. (1975). An Analysis of the Behavior Of A
Class Of Genetic Adaptive System. PhD thesis, Uni-
versity of Mitchigan.
Digalakis, J. G. and Margaritis, K. G. (2002). An exper-
imental study of benchmarking functions for genetic
algorithms. International Journal of Computer Math-
ematics, 79(4):403–416.
Goldberg, D. E. (1989). Genetic Algorithms in Search, Op-
timization, and Machine Learning. Addison-Wesley
Pulishing Company, INC,, Reading, Massachusetts,
US.
Goldberg, D. E. and Deb, K. (1991). A comparative anal-
ysis of selection schemes used in genetic algorithms.
Foundations of genetic algorithms, 1:69–93.
Goldberg, D. E. and Smith, R. E. (1987). Nonstationary
function optimization using genetic algorithms with
dominance and diploidy. In ICGA, pages 59–68.
Grefenstette, J. J. (1986). Optimization of control param-
eters for genetic algorithms. IEEE Transactions on
systems, man, and cybernetics, 16(1):122–128.
Grefenstette, J. J. et al. (1992). Genetic algorithms for
changing environments. In PPSN, volume 2, pages
137–144.
Holland, J. H. (1992). Adaptation in Natural and Artificial
Systems: An Introductory Analysis with Applications
to Biology, Control and Artificial Intelligence. MIT
Press, Cambridge, MA, USA.
Krishnakumar, K. (1990). Micro-genetic algorithms for sta-
tionary and non-stationary function optimization. In
1989 Advances in Intelligent Robotics Systems Con-
ference, pages 289–296. International Society for Op-
tics and Photonics.
Krishnakumar, K. and Bailey, J. (1990). Inertial energy dis-
tribution error control for optimal wind-shear penetra-
tion. Journal of Guidance, Control, and Dynamics,
13(6):944–951.
Leung, Y., Gao, Y., and Xu, Z.-B. (1997). Degree of popula-
tion diversity-a perspective on premature convergence
in genetic algorithms and its markov chain analysis.
IEEE Transactions on Neural Networks, 8(5):1165–
1176.
Mahfoud, S. W. (1995). Niching methods for genetic algo-
rithms. Urbana, 51(95001):62–94.
Mori, N. and Kita, H. (2000). Genetic algorithms for adap-
tation to dynamic environments-a survey. In Industrial
Electronics Society, 2000. IECON 2000. 26th Annual
Confjerence of the IEEE, volume 4, pages 2947–2952.
IEEE.
Mumford, C. L. (2004). Simple population replacement
strategies for a steady-state multi-objective evolution-
ary algorithm. In Genetic and Evolutionary Computa-
tion Conference, pages 1389–1400. Springer.
Whitely, D. (1989). Optimizing neural networks using
faster, more accurate genetic search. Proc. of ICGA-
89.
Yang, S. (2008). Genetic algorithms with memory-and
elitism-based immigrants in dynamic environments.
Evolutionary Computation, 16(3):385–416.
