
Muscle Fiber Function during Rapid Movement
based Solely on Kinesthesia
K. Ogiso
1
, K. Hirose
1
, M. Takenaka
1
, D. Nagaoka
1
and M. Tokui
2
1
Department of Education, Kogakkan University, 1704 Kodakujimoto, Ise, Japan
2
Department of Sport Science, Kyushu Kyoritsu University, 1-8 Jiyugaoka, Yahatanishi-ku, Kitakyusyu, Japan
Keywords: Maximum Knee Extension, Vastus Lateralis Muscle, Pennation Angle, Aponeurosis, Reaction Time, Pre-
Activity.
Abstract: This study was designed to examine function of the vastus lateralis muscle (VL) fibers during maximal
voluntary contraction (MVC) in knee extension which was exerted based solely on the kinesthesia acquired
from repeating the MVC movements. Fifteen men performed 10 consecutive isokinetic knee extensions
comprising 7 passive contractions and 3 MVCs, which was repeated for 7 sets. In the first 3 sets, subjects
were instructed to perform MVCs immediately a light cue appeared when the leg reached 60 deg knee joint
angle in the 3rd, 6th, and 9th extensions; in the next 4 sets, subjects tried to maintain the timing of MVC
repetitions without the light cue. VL electromyographic activity was monitored. The point where a fascicle
arose from the deep aponeurosis and the pennation angle were measured on VL ultrasonic images. Subjects
classified their MVC performance (force and timing) into 5 grades after each set. Based solely on
kinesthesia (without the light cue), the VL fibers contracted tightly to a point where the fascicle arises from
the deep aponeurosis, and it appeared to compensate for a delay in reaction time to start MVC. However, the
subject’s self-evaluation remained unchanged despite the changes in muscle behavior during MVC. In the
4th set only, when the light cue was not used for the first time, did their self-evaluation tend to decrease and
VL pre-activity was significantly increased. These results suggest that kinesthesia does not always
correspond to actual muscle activity.
1 INTRODUCTION
We perform physical movement based on
kinesthesia to achieve a given motor task. However,
the outcome is not always what we hoped for. If a
delay occurs between kinesthesia and our actual
movement, a large force must be exerted in a quite
short time to bridge them.
In the pennate muscles, contraction is performed
by shortening the muscle fibers (Fukunaga et al.,
1997; Hawkins and Bey, 1997) and increasing the
pennation angle (Ichinose et al., 1997; Maganaris
and Baltzopoulos, 1999), which rotate the joints
throughout the tendinous tissues elongated by the
muscle. However, the interaction between such
shortening and the degree of the pennation angle is
not constant and varies between low- and high-force
contractions (Azizi et al., 2008), suggesting that
sudden modulation of the muscle force is likely to
induce irregular muscle fiber behavior. In fact, the
muscle fibers have been reported to contract more
tightly near the deep aponeurosis when the timing to
exert force was unexpectedly changed (Hirose et al.,
2013).
Muscle contraction which is performed based
solely on the kinesthesia may not basically give
agreement with muscle fiber behavior which is
induced based on a lot of information. Especially in
rapid movements, a feed-forward control of the
central nervous system which is likened to learned
anticipatory responses to known cues plays an
important role to control the rapid motions because a
feedback component is very slow. This suggests that
the function of the muscle fibers may vary
depending on whether information has been received
to start the muscle contraction.
A clinical report revealed that most muscle strain
injuries occur at or near the myotendinous junction
during high-intensity or explosive voluntary
movements such as sprint and quick turn (Okuwaki,
2009). If the muscle fibers are strong and shorten
unexpectedly with an inappropriate timing, it may
11
Ogiso K., Hirose K., Takenaka M., Nagaoka D. and Tokui M..
Muscle Fiber Function during Rapid Movement based Solely on Kinesthesia.
DOI: 10.5220/0005142900110016
In Proceedings of the 2nd International Congress on Sports Sciences Research and Technology Support (icSPORTS-2014), pages 11-16
ISBN: 978-989-758-057-4
Copyright
c
2014 SCITEPRESS (Science and Technology Publications, Lda.)

induce irregular muscle fiber behavior and lead to
muscle strain injuries with higher probability.
Therefore, we designed this study to examine the
characteristics of muscle fiber functions during
maximal voluntary contraction (MVC) in knee
extension which was exerted based solely on the
kinesthesia acquired from repeating the movement at
a constant timing indicated by a light cue.
2 METHODS
2.1 Subjects
Fifteen men (age, 21.9±1.1 years, height 172.6±9.1
cm, weight 70.5±10.7 kg) participated in this study.
All subjects were in good health, with no orthopedic
or neuromuscular abnormalities. Subjects were fully
informed of the nature and possible consequences of
the study before providing written informed consent.
The experiments were conducted in accordance with
the Declaration of Helsinki. Approval was obtained
from the Ethics Committee of Kogakkan University.
2.2 Measurement Procedure
The subjects completed a warm-up consisting of
jogging and dynamic stretching for 10 min. Then
they were placed in a comfortable, upright, seated
position on an isokinetic dynamometer chair, and the
dynamometer fulcrum was aligned with the axis of
rotation of the left knee joint (Biodex-System 4,
Biodex Medical Systems, New York, USA).
Subjects were secured using shin, thigh, pelvic, and
torso stabilization straps to minimize extraneous
body movements and were asked to fold their arms
across their chest during the experiment. After
correcting for the effects of gravity, several sub-
maximal or maximal knee extensions were repeated
in a second warm-up period. The knee movement
range of motion was from 0 deg extension to 90 deg
flexion and was tested at 90 deg/s (0 deg = straight
leg).
After the subjects familiarized themselves with
the experimental apparatus and procedure by
performing knee extensions at MVC several times,
they performed 10 consecutive isokinetic knee
extensions comprising 7 passive contractions and 3
MVCs. All MVCs were performed during the
passive isokinetic knee extensions and were
followed by passive knee flexions. The 10
consecutive isokinetic knee extensions were
repeated for 7 sets with 2-min intervals. A photo
beam unit consisting of light-emitting and light-
receiving devices was set up on either side of the left
shank such that an LED placed in front of the
subject could be switched on, to act as a light cue to
start MVC, when the shin moved through the beam
at 60 deg knee joint angle during knee extension in
the first 3 sets. Immediately before reaching 60 deg
knee joint angle, the angular velocity had reached a
constant velocity of 90 deg/s.
In the first 3 sets, subjects were asked to relax
their muscles and exert an MVC immediately they
saw the light cue during the 3rd, 6th, and 9th
repetitions. They were informed beforehand of the
timing when the light cue would come on. In the
next 4 sets, based solely on the kinesthesia acquired
during the first 3 sets, the subjects tried to exert
MVCs at the same timing as in the first 3 sets but
without the light cue. After each set, the subjects
classified the force and timing of their MVC
performance into 5 grades (1 = very poor; 2 = poor;
3 = average; 4 = good; 5 = very good).
2.3 Data Collection
The knee joint torque (KJT) exerted at MVC was
measured with data on the knee joint angle (KJA).
Electromyographic (EMG) activity was recorded
from the vastus lateralis muscle (VL) in the left leg,
using Ag/AgCl bipolar surface electrodes (diameter,
10 mm; inter-electrode distance, 20 mm; TELEmyo
DTS, Noraxon, Scottsdale, USA). EMG signals
were amplified. A/D was converted at a sampling
rate of 1.5 kHz and transmitted to a computer along
with data on KJT, KJA, and timing of the light cue.
Longitudinal sectional images of the VL were
obtained at 37 Hz using a real-time B-mode
ultrasound apparatus (Prosound α7, Hitachi Aloka
Medical, Tokyo, Japan). A linear array probe with a
scanning frequency of 7.5 MHz and a scanning
length of 60 mm was fixed with a sponge and an
elastic bandage over the VL at one-quarter from the
distal end of the estimated muscle length. The
ultrasonic images with the timing of the light cue
were transmitted to a computer and recorded onto
Blu-ray discs.
2.4 Data Processing
The EMG signals were full-wave rectified. The
following 4 reaction time characteristics were
measured: time between the light cue and onset of
EMG activity (premotor reaction time; PMT), time
between the onset of EMG activity and onset of KJT
(electromechanical delay; EMD), time between the
light cue and onset of KJT (total reaction time; TRT),
icSPORTS2014-InternationalCongressonSportSciencesResearchandTechnologySupport
12
and time between the light cue and peak KJT
(movement time; MTPT). Average EMG (aEMG)
was calculated over 100 ms before the light cue as a
measure of pre-activity.
For each ultrasonic image, the following 6 points
were digitized and converted to real coordinates:
point (P) where a fascicle arises from the deep
aponeurosis; point (S) where a perpendicular line
from P intersects the superficial aponeurosis; 2
points (F
5
and F
10
) on the fascicle 5 mm and 10 mm
horizontally from P, respectively; and 2 points (D
5
and D
10
) on the deep aponeurosis 5 mm and 10 mm
horizontally from P, respectively (Fig.1). Digitizing
the 6 points was repeated 3 times for each image and
the coordinates were averaged. The interior angles
∠F
5
PD
5
and ∠F
10
PD
10
were calculated as 2 types
of pennation angle (PA
5
and PA
10
, respectively). The
distance between points P and S was taken as the
muscle thickness.
2.5 Statistics
Data are presented as the means ± SD. One-way
analysis of variance (ANOVA) was used to analyze
the differences in aEMGs, reaction times and 5-
grade evaluations of MVC performance. To test for
the effects of kinesthesia on the behaviour of the
muscle fibers, two-way ANOVAs (factors: set vs.
pennation angle) for repeated measurements were
performed. Fisher’s post hoc comparison was
performed when significance was found. The
probability level accepted for statistical significance
was p<0.05.
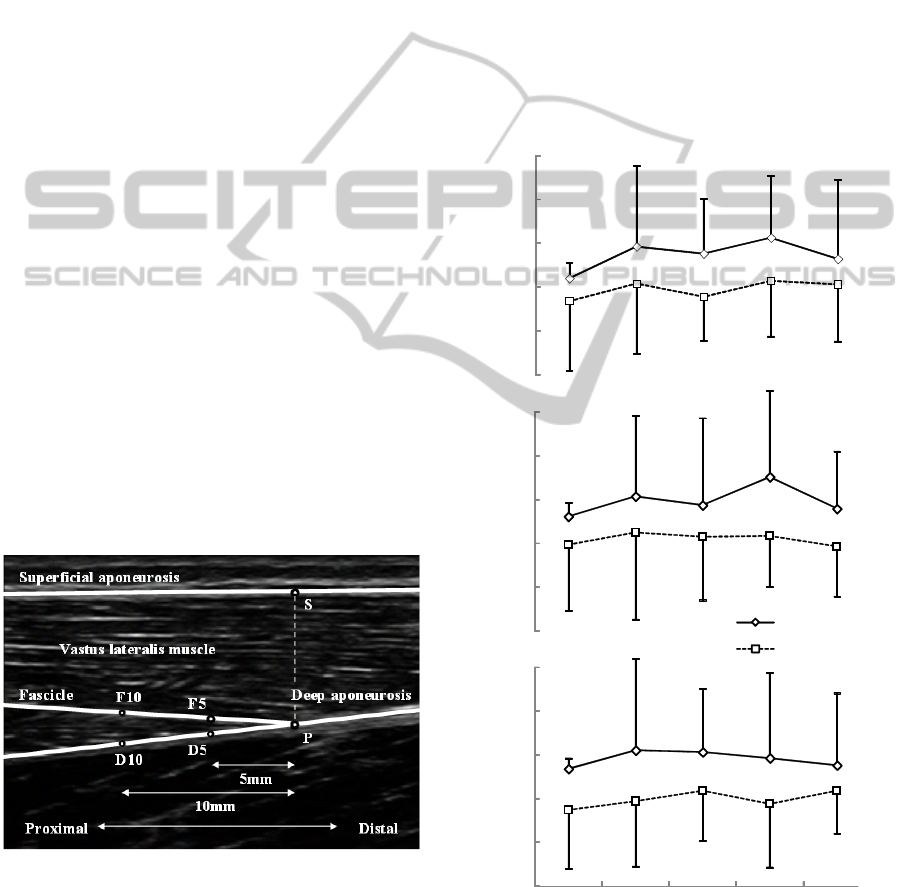
Figure 1: Analysis of ultrasonic image of the vastus
lateralis muscle. Six points were digitized. Two types of
pennation angle (∠F
5
PD
5
and ∠F
10
PD
10
, respectively)
and the muscle thickness (the distance between points P
and S) were calculated.
3 RESULTS
Increases in PA
5
from rest to peak KJT tended to be
larger in the 4th to 7th sets without the light cue than
in the 1st to 3rd sets with the light cue (Fig.2).
Standard deviations of the increase in PA
5
among the
3rd, 6th, and 9th repetitions in each set were
significantly larger in the 4th to 7th sets than in the
1st to 3rd sets (Fig.3). On the other hand, the
influence of the light cue on PA
10
was smaller than
that on PA
5
. The moving speed of P became more
unstable during MVC in the 4th to 7th sets without
the light cue. No significant differences were
observed in muscle thickness in each set.
Figure 2: Increase in pennation angle
from rest to peak
knee joint torque.
5
10
15
20
25
30
∠
F
5
P D
5
∠
F
10
P D
10
5
10
15
20
25
30
5
10
15
20
25
30
1st-3rd 4th 5th 6th 7th
Pennation angle (deg)
3rd repetition
6th
9th
Set
MuscleFiberFunctionduringRapidMovementbasedSolelyonKinesthesia
13
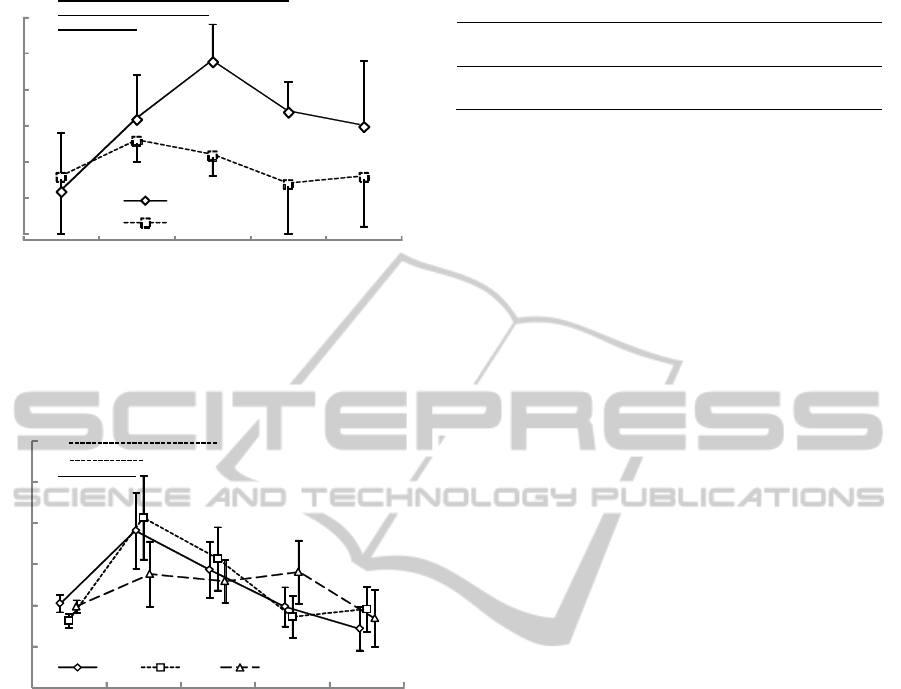
Figure 3: Standard deviation of the increase in pennation
angle from rest to peak knee joint torque. Asterisks
indicate significant differences between sets (* p<0.05, **
p<0.01).
Figure 4: Average electromyography 100ms before flexion
to 60deg knee joint angle (pre-activity). Asterisks indicate
significant differences between sets (p<0.05).
No significant differences were noted in the
increasing rate of KJT between the 1st to 3rd sets
and the 4th to 7th sets. The standard deviations in
the 3rd, 6th, and 9th repetitions increased as the set
was repeated and were significantly larger in the 7th
set than in the 1st to 3rd sets.
PMT tended to be longer in the 4th to 8th sets
than in the 1st to 3rd sets. It increased from about
40ms in the 1
st
to 3
rd
sets to about 100ms in the 4
th
to
8
th
sets. Standard deviations of the PMT in the 3rd,
6th, and 9th repetitions in each set tended to be
increased in the 4th to 7th sets than in the 1st to 3rd
sets. A significant difference was observed between
the 4th set and the 1st to 3rd sets. However, no
significant differences were noted in EMD in each
set regardless of the presence of a light cue. TRT,
Table 1: Self-evaluation of MVC on a 5-point scale.
Set
1st to 3rd
4th 5th 6th 7th
Force
2.9±0.5 2.6±0.7 2.8±0.4 2.8±0.8 3.0±0.7
Timing
3.1±0.7 2.6±0.5 3.2±0.4 3.0±0.7 2.8±0.8
which is the sum of PMT and EMD, tended to be
similar to the PMT.
In terms of pre-activity, some significant
differences in aEMG were observed between the 1st
to 3rd sets and the 4th or 5th set (Fig.4). The
differences seen immediately after the light cue had
disappeared decreased as the set or repetition was
repeated, and the aEMG eventually tended to be
equal to or smaller than that in the 1st to 3rd sets.
No significant differences were observed in the
self-evaluation of MVC performance in each set.
However, the scores for both force and timing
tended to decrease only in the 4th set, which was the
first set performed without the light cue (Table 1).
4 DISCUSSION
Loss of the light cue to start MVC altered the
behavior of the muscle fibers during contraction.
Exerting MVC based solely on kinesthesia without
the light cue made the moving velocity of P unstable
and increased both PA near the deep aponeurosis
and PMT. However, the subjects themselves could
not perceive these changes; self-evaluation of the
force and timing of MVC repetitions remained
unchanged regardless of the presence of a start
(light) cue. This concurs with the findings of a
previous study which found that subjects also did
not perceive differences in the function of the
muscles during MVC repeated at a constant timing
when the timing of the muscle contraction was
unexpectedly changed (Hirose et al., 2013). This
suggests that kinesthesia does not always correspond
to the actual movement performed.
The subject’s self-evaluations tended to decrease
slightly in the 4th set only, which was the first set
without the light cue. This indicates that alterations
in the muscle behavior can be vaguely perceived
immediately after the condition for exerting MVC
was changed. Indeed, repetitive exercise has been
reported to improve the reproducibility of the
movement to adjust force and position to target
levels (Kaneko et al., 2009), and motor experience
has been reported to improve somatosensory
functions (Hayami et al., 2008). These findings
suggest that sufficient time is needed to perceive the
0
1
2
3
1st-3rd 4th 5th 6th 7th
∠
F
5
P D
5
∠
F
10
P D
10
Set
(deg)
*
**
*
0
20
40
60
80
100
120
1st-3rd 4th 5th 6th 7th
3
rd
6
th
9
th
repetition
Set
Average EMG (µV)
*
*
*
icSPORTS2014-InternationalCongressonSportSciencesResearchandTechnologySupport
14

muscle behavior and that it is difficult to maintain
kinesthesia acquired only over a brief period.
In the sets without the light cue in this study,
significantly larger pre-activity was observed before
the onset of MVC. This implies that the muscle was
preparing to establish the timing to exert MVC. It is
known that pre-activity increases so that fatigue-
induced declines in performance do not deteriorate
further (Horita et al., 1999). Therefore, pre-activity
is likely to be a preparatory condition for generating
as large an MVC as possible at an appropriate timing
based solely on kinesthesia, which may explain the
lower self-evaluations of MVC in the 4th set. Pre-
activity gradually decreased after the 4th set and
became closer to the smaller values seen in the 1st to
3rd sets, although the standard deviations of PMT
and PA
5
in the 3rd, 6th, and 9th repetitions were
increased. This suggests a process whereby the
accuracy of movement based on kinesthesia is
decreased due to the disappearance of kinesthetic
information.
Behavior of the muscle fibers during MVC based
solely on kinesthesia was characterized by a large
and unstable PA
5
. The muscle fibers contracted
tightly to a point where the fascicle arises from the
deep aponeurosis, which appeared to compensate for
the delay in the reaction time to start MVC, and
similar muscle fiber behavior has been observed
when the timing to exert MVC was unexpectedly
changed (Hirose et al., 2013). Since muscle force is
transmitted through connective tissues to
neighboring muscles (Huijing, 2003; Sandercock
and Haas, 2009), the behaviour of the muscle fibers
near the deep aponeurosis might result from the
force transmission, and it might be influenced also
by pressure put with a probe fixed over the muscle.
Further studies are needed to elucidate the
mechanism causing strong muscle fiber contractions
near the deep aponeurosis.
Muscle strain injury was reported to occur at or
near the myotendinous junction in frog
myotendinous units when the muscle was strained
(Tidball et al., 1993). Similarly, in human muscles,
muscle failure is created by combining a large force
with substantial stretch near the aponeurosis
(Garrett, 1990), and a clinical study reported most
muscle strain injuries occurring at or near the
myotendinous junction during high-intensity or
explosive voluntary movements (Okuwaki, 2009). If
the muscle fibers are strong and shorten
unexpectedly near the deep aponeurosis with an
inappropriate timing during stretch, the contraction
will increase the inhomogeneous strain on the
aponeurosis (Zuurbier et al., 1994; Kinugasa et al.,
2008), which may cause a muscle strain injury at or
near the myotendinous junction.
In conclusion, behavior of the muscle fiber
during MVC which was exerted based solely on
kinesthesia without the light cue was characterized
by stronger and more unstable contraction near the
deep aponeurosis with longer premotor reaction time
and larger pre-activity. However, the subjects could
not perceive these changes. Such irregular muscle
fiber behavior may be related to a mechanism of
muscle injury.
REFERENCES
Azizi, E., Brainerd, E.L., Roberts, T.J., 2008. Variable
gearing in pinnate muscles. PNAS 105: 1745-1750.
Fukunaga, T., Ichinose, Y., Ito, M., Kawakami, Y.,
Fukashiro, S., 1997. Determination of fascicle length
and pennation in a contracting human muscle in vivo.
J Appl Physiol 82: 354-358.
Garrett, W.E., Jr., 1990. Muscle strain injuries: clinical
and basic aspects. Med Sci Sports Exerc 22: 436-443.
Hawkins, D., Bey, M., 1997. Muscle and tendon force-
length properties and their interactions in vivo. J
Biomech 30: 63-70.
Hayami, T., Kaneko, F., Kizuka, T., 2008. Differences in
the Function of Somatosensory-Motor Integration
depend on the Motor Experiences. SOBIM 19: 47-56.
Hirose, K., Tarodachi, N., Tsutsumi, M., Ogiso, K., 2013.
Timing of muscle contraction influences function of
muscle fibers. In Book of Abstract, 18
th
annual
Congress of the European College of Sport Science.
SporTools.
Horita, T., Komi, P.V., Nicol, C., Kyrolainen, H., 1999.
Effect of exhausting stretch-shortening cycle exercise
on the time course of mechanical behaviour in the
drop jump: possible role of muscle damage. Eur J
Appl Physiol 79: 160-167.
Huijing, PA., 2003. Muscular Force Transmission
Necessitates a Multilevel Integrative Approach to the
Analysis of Function of Skeletal Muscle. Exerc. Sport
Sci. Rev 31(4): 167-175.
Ichinose, Y., Kawakami, Y., Ito, M., Fukunaga, T., 1997.
Estimation of active force-length characteristics of
human vastus lateralis muscle. Acta Anat 159: 78-83.
Kaneko, F., Hayami, T., Yokoi, T., Kizuka, T., 2009.
Research on Examination Indicator of Motor
Performance Corresponding to the Active Kinesthetic
Perception: from the Viewpoint of Detection of
Repetitive Practice Effect. Physical Therapy Japan
36(1): 9-17.
Kinugasa, R., Shin, D., Yamauchi, J., Mishra, C., Hodgson,
J.A., Edgerton, V.R., Sinha, S., 2008. Phase-contrast
MRI reveals mechanical behavior of superficial and
deep aponeuroses in human medial gastrocnemius
during isometric contraction. J Appl Physiol 105:
1312–1320.
MuscleFiberFunctionduringRapidMovementbasedSolelyonKinesthesia
15

Maganaris, CN., Baltzopoulos, V., 1999. Predictability of
in vivo changes in pennation angle of human tibialis
anterior muscle from rest to maximum isometric
dorsiflexion. Eur J Appl Physiol 79: 294-297.
Okuwaki, T., 2009. Muscle strains of top-level athletes in
Japan. J Japan Soc Clin Sports Med 17: 497-505.
Tidball, J.G., Salem, G., Zernicke, R., 1993, Site and
mechanical conditions for failure of skeletal muscle in
experimental strain injuries. J Appl Physiol 74: 1280-
1286.
Zuurbier, C.J., Everard, A.J., van der Wees. P., Huijing,
P.A., 1994. Length-force characteristics of the
aponeurosis in the passive and active muscle condition
and in the isolated condition. J Biomech 27: 445–453.
icSPORTS2014-InternationalCongressonSportSciencesResearchandTechnologySupport
16
