
Modeling Embedded Interpersonal and Multiagent Coordination
Michael J. Richardson
1
, Rachel W. Kallen
1
, Patrick Nalepka
1
, Steven J. Harrison
2
, Maurice Lamb
1
,
Anthony Chemero
1
, Elliot Saltzman
3
and R. C. Schmidt
4
1
Center for Cognition, Action and Perception, University of Cincinnati, Cincinnati, OH, U.S.A.
2
School of Health, Physical Education and Recreation, University of Nebraska Omaha, Omaha, NE, U.S.A.
3
Department of Physical Therapy and Athletic Training, Sargent College of Health and Rehabilitation Sciences,
Boston University, Boston, MA, U.S.A.
4
Department of Psychology, College of the Holy Cross, Worcester, MA, U.S.A.
Keywords: Multiagent Systems, Social Coordination, Task Dynamics, Complex Systems, Self-organization.
Abstract: Interpersonal or multiagent coordination is a common part of everyday human activity. Identifying the
dynamic processes that shape and constrain the complex, time-evolving patterns of multiagent behavioral
coordination often requires the development of dynamical models to test hypotheses and motivate future
research questions. Here we review a task dynamic framework for modeling multiagent behavior and
illustrate the application of this framework using two examples. With an emphasis on synergistic self-
organization, we demonstrate how the behavioral coordination that characterizes many social activities
emerges naturally from the physical, informational, and biomechanical constraints and couplings that exist
between two or more environmentally embedded and mutually responsive individuals.
1 INTRODUCTION
Many of the everyday movements and actions that
individuals perform are accomplished in a social
setting and are coordinated with the actions of other
individuals. Such behavioral activity is sometimes
deliberate and conscious, like when two people are
moving a large piece of furniture together, or
spontaneous and automatic, such as when two
people avoid bumping into one another when
walking on a busy sidewalk. It can involve the
synchronous coordination of rhythmic movements,
such as when individuals synchronize their legs
movements while walking and talking, or the
asynchronous coordination of complementary or
discrete actions, such as when loading a dishwasher
together. Finally, multiagent coordination tasks often
involve a nested structure of sub-actions, with the
varied sequencing of these actions making up the
complex syntax of everyday social activity.
The varied complexity of social and multiagent
coordination, however, belies the seemingly
effortless and robust manner with which human (and
many animal) agents are able to perform these
activities. Indeed, two or more human agents can
often spontaneously coordinate their movements and
actions in highly novel task contexts with little to no
prior experience or learning. This apparent context
insensitivity has led many researchers to focus on
trying to identify the invariant set of internal
(mental) neurocognitive and perceptual-motor
mechanisms that might support effective multiagent
coordination across a wide range of task goals and
situations (Graf, et al., 2009; Newman-Norland et
al., 2007; Vesper et al., 2010). It is becoming
increasingly clear, however, that that the CNS
cannot “do it all”, and that there is more to social
action and coordinated multiagent activity than
simply neurocognitive processes alone (Coey et al.,
2012; Eiler et al., 2013; Knoblich, et al., 2011;
Schmidt et al., 2011). This is particularly true for
complex multiagent tasks that require individuals to
continuously adapt their behavior to each other and
to a dynamic task environment, where adopting a
purely neurocognitive approach requires increasing
appeals to ever higher levels of ungrounded,
representational complexity, as well as unsustainable
levels of executive control (Chemero, 2009; Schmidt
et al., 1990; 2011).
So what other processes or mechanisms might
Richardson, M., Kallen, R., Nalepka, P., Harrison, S., Lamb, M., Chemero, A., Saltzman, E. and Schmidt, R.
Modeling Embedded Interpersonal and Multiagent Coordination.
In Proceedings of the 1st International Conference on Complex Information Systems (COMPLEXIS 2016), pages 155-164
ISBN: 978-989-758-181-6
Copyright
c
2016 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved
155
operate to shape and constrain the complex time-
evolving patterns of behavioral coordination that
characterize everyday interpersonal or multiagent
activity? Here, we propose that many of the stable
patterns of interpersonal and multiagent coordination
that occur during social activity lawfully emerge
from the physical and informational constraints and
couplings that exist between two or more
environmentally embedded and mutually responsive
individuals (Marsh et al., 2009; Richardson and
Kallen, 2015; Schmidt et al., 1990; Schmidt et al.,
2011). Of particular relevance here is the resultant
implication that context dependent multiagent
coordination can be understood and modeled as a
coupled nonlinear dynamical system. Accordingly,
the aim of the current paper is to provide a general
overview of how one can employ coupled nonlinear
dynamical models to capture and explain the robust,
time-evolving structure of Embedded Multi-Agent
Dynamics (EMAD). Drawing heavily on the task
(Saltzman and Kelso, 1987) and behavioral dynamic
(Warren, 2006) approaches to human behavior, we
first briefly define a conceptual framework for
modeling EMAD. We then offer paradigmatic
examples illustrating how this framework can be
employed to formalize low-dimensional, coupled,
nonlinear differential equation models of
coordinated interpersonal and multiagent behavior.
2 MODELING FRAMEWORK
Consistent with more general nonlinear dynamics
and complex systems approaches to human behavior
(for reviews see e.g., Kelso, 1995; Richardson et al.,
2014; Thelen and Smith, 1994; Warren, 2006), and
building on the dynamical interpersonal coordination
research of Schmidt and colleagues (e.g., Schmidt et
al, 1990; Schmidt and O’Brien, 1997; Schmidt and
Turvey, 1994), the framework for modeling EMAD
detailed here emphasizes self-organization and
contextual emergence. Accordingly, the stable
organization of multiagent behavior is not captured
by means of a centralized control structure or
neurocognitive mechanism, but rather is modeled as
an emergent, a posteriori consequence of the
distributed interaction of physical processes,
informational and neurological couplings, and
contextual constraint. Accordingly, stable patterns of
behavioral coordination correspond to functional
(re)organizations of a multiagent-environment
system (e.g., limbs, movements of different actors,
objects within a social task environment), one that is
temporarily constrained to act as a single coherent
unit or synergy, formed and destroyed in response to
changing task goals and perceived action
possibilities (Riley, et al., 2011).
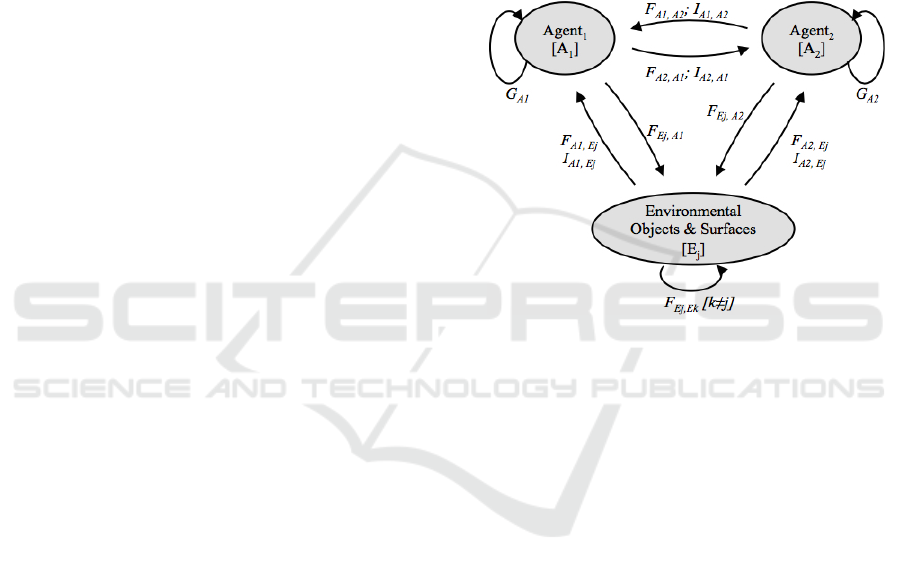
More formally, environmentally embedded
interpersonal or multiagent coordination emerges
from the activity of two or more agents, A
i
(where i
= 1, 2, … n agents) in a changing task environment,
E, via the detection of information, I, and the
reciprocal and mutually constraining modulation of
behavioral action and environmental events by the
physical forces, F, exerted in the environment by the
agents or by other environmental objects or surfaces
(or by both).
Figure 1: Illustration of a task dynamic framework for
modeling Embedded Multi-agent Dynamics (EMAD). See
text for details.
A two-agent (interpersonal) illustration of this
multiagent-environment modeling framework is
illustrated as a system graph in Fig. 1. Here, each
agent, i, and task relevant environmental objects and
surfaces, j, are represented as nodes, with the
different nodes linked via mechanical forces, F, and
information, I. Note that an agent’s behavior is also
a function, G, of intrinsic dynamical or self-
referential processes. Similarly, environmental
objects or surfaces can also be influenced via
physical, F, object-object and object-surface
interactions.
This graph representation defines the “bottom-
up-all-the-way-down” nature of EMAD, where the
general aim of modeling such behavior is to
formalize the simplest set of functional relations that
give rise to observed macroscopic behavioral order.
Different task systems will of course entail different
graphs, each needing to be formalized into a specific
system of dynamical equations. This is best achieved
by defining a functional description of the
multiagent task dynamics in terms of an abstract task
space. Similar to the intrapersonal task dynamic
COMPLEXIS 2016 - 1st International Conference on Complex Information Systems
156
formalism of Saltzman and Kelso (1987), this
includes appropriately defining (i) the task goal in
terms of the relevant terminal objective, (ii) the
minimal number of task dimensions (i.e., relevant
task axes or variables), which correspond to the
active degrees-of-freedom of the task’s end-effector,
and (iii) the appropriate task dynamics (equations of
motion) for each task dimension; equations that
should qualitatively capture the movement or action
trajectories that are afforded along each task axis of
the terminal objective.
Admittedly, defining the relevant task
dimensions and appropriate equations of motion for
a specific multiagent task is not always easy and can
require an extensive amount of empirical research
and theorizing, not to mention a considerable
amount of trial-and-error. It is with this mind that we
have chosen to review a range of simple, yet
ubiquitous joint-action tasks and behaviors in order
to best illustrate this modeling process and how task
dynamic models (specifically with respect to
differential equation models) can be formulated to
capture and understand the interactive and self-
organizing processes that determine coordinated
multiagent behavior.
3 RHYTHMIC COLLISION
AVOIDANCE
As a first example of how to model the task
dynamics of goal directed multiagent coordination,
we consider the interpersonal rhythmic collision
avoidance task investigated by Richardson et al.,
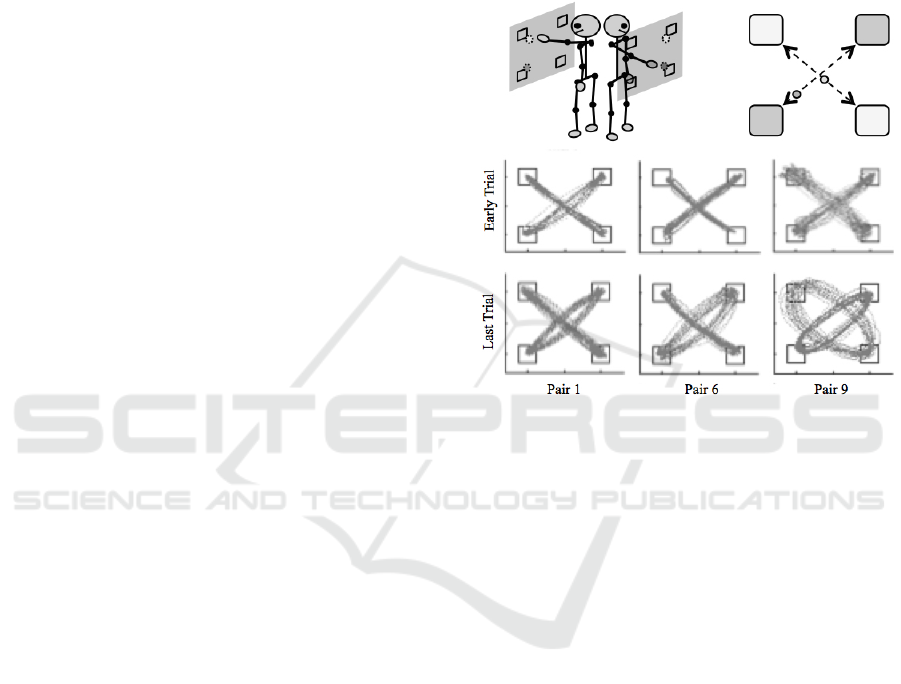
(2015). For this task, pairs of naive participants were
instructed to continuously move a visual, computer
cursor (a small red dot) back and forth between
different sets of square target locations, positioned at
diagonally opposite corners of a 50” computer
monitor. Each participant in a pair stood facing their
own computer monitor, which displayed the real-
time motion of the participant’s own cursor, as well
as the motion of their co-participant’s cursor.
Participants controlled their respective computer
cursor using a motion-tracking sensor (see Fig. 2
top). One participant was instructed to continuously
move their cursor between the bottom-left and top-
right targets while the other participant was
instructed to continuously move their cursor
between the bottom-right and top-left targets. Most
importantly, participants were instructed to produce
these continuous cursor motions without colliding
into one another.
In this task, participant pairs were faced with a
conflict between the natural tendency to synchronize
straight-line movement trajectories between the
targets and the fact that such synchronization would
result in a collision. Thus, the research question
being investigated was what stable pattern(s) of
movement coordination would emerge to ensure task
success?
Figure 2: Illustration of the experimental setup (top left)
and task display (top right) employed in the Richardson et
al (2015) collision avoidance task. (bottom) Prototypical
examples of the movement trajectories exhibited by pairs.
Although pairs initially tried to adopt relatively
straight line trajectories between the target locations,
the results revealed that greater than 90% of pairs
quickly converged (by the 3
rd
or 4
th
trial) onto a
stable task solution that involved complementary
task roles, with one participant adopting a more
straight-line trajectory between targets and the other
participant adopting a more elliptical trajectory
between targets (see Fig. 2, bottom). Moreover, the
participant who adopted the more elliptical
trajectory consistently lagged behind the participant
who adopted the more straight-line trajectory by an
average of approximately 10° to 30°. Richardson et
al., (2015) hypothesized that these complementary
behavioral dynamics were the result of a functional
asymmetry in the repulsive coupling that prevented
collisions, one that not only resulted in an inter-
participant asymmetry in the ellipticality of the
movement trajectories, but also simultaneously
allowed participants to synchronize their between
target movements at a phase lag (further increasing
the margin of safety). Of particular relevance to our
discussion here, is that Richardson et al. were able to
Modeling Embedded Interpersonal and Multiagent Coordination
157

test this hypothesis by developing a task dynamic
model of the interpersonal collision avoidance task.
Figure 3: (a) Illustration of the abstract task space axis for
the collision avoidance model developed by Richardson, et
al., (2015). (b) The task space orientated in the target goal
space for a pair of coupled individuals. See text for more
details.
Motived by the same task dynamic framework
described above, Richardson, et al., (2015)
developed this model by defining the terminal
objective of each actor’s task goal to be a rhythmic
motion of the end-effector (i.e., stimulus/end-
effector/hand, modeled as a point mass) between
two targets within a planar (two-dimensional) task
space (see Fig. 3a). In this task space, the x-axis
corresponded to the axis of instructed direction of
oscillation, and was provided with limit cycle
(oscillatory) dynamics. The orthogonal task axis, y,
corresponded to deviations away from the x-axis.
Given that an actor must minimize these deviations
with respect to achieving the instructed task goal,
Richardson et al. defined the y-axis with simple
point-attractor (damped mass-spring) dynamics.
Assuming a point mass of 1 (for simplicity), the
functional defined task space was then specified as:
−
+
+
=0
+
+
=0
(1a)
for actor 1 and
−
+
+
=0
+
+
=0
(1b)
for actor 2. Here,
and
,
,
and
,
and
correspond to the position, velocity and
acceleration of each actor’s end effector (i.e.,
hand/cursor) along each task axis, respectively. b
xi
and b
yi
are the damping coefficients for axis x
i
and y
i
,
respectively, k
xi
and k
yi
are stiffness coefficients for
axis x
i
and y
i
, respectively, and (
) is the van
der Pol function for axis x
i
.
To capture the natural entrainment observed
between co-actors, Richardson and colleagues then
added an attractive coupling function to each
system, similar to what is typically employed for
modeling generalized rhythmic (inphase)
coordination. That is, they added the diffusive
coupling functions
(
−
)
(2a)
and
(
−
)
(2b)
to the equations that define each actor’s instructed
axes of motion, which resulted in the system
−
+
+
=
(
−
)
+
+
=0
−
+
+
=
(
−
)
+
+
=0
(3)
It is important to note at this point that Eq. (3)
provides an idealized model of inphase interpersonal
rhythmic coordination and can be adapted to model
the inphase coordination of any two end effectors or
point masses (e.g., finger tips; hands, feet, actors)
irrespective of the angular orientation of the
instructed motion axis. Moreover, other behavioral
patterns, such as antiphase coordination and the
bistable nature of rhythmic coordination performed
in the same direction/plane of motion can also be
captured using alternative coupling functions, e.g.,
as previously identified by Haken et al., (1985) (also
see e.g., Dumas et al., 2014; Kelso, 1995; Schmidt
and Richardson, 2008)
Based on the behavior of participants at the
beginning of the experiment (see Fig. 2),
Richardson, et al. (2015) began with the assumption
that pairs were initially constrained by the dynamics
of Eq. (3). The question then became what minimal
changes in the structure of Eq. (3) were required to
produce behavior qualitatively similar to that
produced by pairs by the end of the experiment?
Given that the task instructions were to ‘avoid
bumping or colliding into each other’, the simplest
modification was to add a repelling coupling force
that acted on each participant’s end-effector to
ensure they were repelled from each other. This was
accomplished using the repeller functions
(
+
)
|
|
(4a)
(
−
)
|
|
(4b)
for the primary task axes, x
1
and x
2,
respectively, and
(
−
)
|
|
(4c)
(
+
)
|
|
(4d)
COMPLEXIS 2016 - 1st International Conference on Complex Information Systems
158

for the secondary task axes, y
1
and y
2,
respectively.
In short, these repeller functions push the two
participants’ end-effectors away from each other, at
a strength determined by the parameters, γ
i
, and
inter-cursor distance, i.e., the exponential terms in
Eq. 4 mean that larger (smaller) distances between
the participants’ cursors result in weaker (stronger)
repelling forces (see Richardson et al., 2015 for
more details about how these coupling functions
were derived). Essentially, if γ
i
in participant-i’s (x,
y) system is set to zero, the effect of the repeller
coupling is null and a straight-line trajectory will be
created that is aligned along the participant’s limit-
cycle axis. However, if γ
i
is set to a value greater
than zero, the repeller coupling adds a force along
both task-axes of participant-i, resulting not only in
greater ellipticality (due to forces added along the
point-attractor task axis), but also in a phase lag
relative to participant-j when γ
i
is greater than γ
j
(due
to forces added along the limit-cycle task axis).
The complete task dynamic model derived by
Richardson et al., (2015) can therefore be written as
−
+
+
=
(
−
)–
(
+
)
|
|
+
+
=
(
−
)
|
|
−
+
+
=
(
−
)
+
(
−
)
|
|
+
+
=−
(
+
)
|
|
(5)
Importantly, this system was not only able to
successfully capture the stable asymmetric task
solution adopted by participants—namely,
asymmetric movement trajectories and phase-lagged
rhythmic synchronization—but it could also capture
the different types of successful and unsuccessful
movement solutions adopted by participants pairs
throughout the course of the experiment.
Essentially, there are three qualitative types of
movement trajectories that can be generated from
Eq. (5). These are displayed in Fig. 4 and depend on
the magnitudes of γ
1
and γ
2
and the degree that γ
1
≠
γ
2
. First, if γ
1
= γ
2
= 0, then no motion is created
along
or
(i.e.
=
= 0), which makes the
behavior of Eq. (5) equivalent to the behavior of Eq.
(3). This corresponds to straight-line inphase
coordination (Fig. 4 left) and as noted previously
was what most participants in the Richardson et al
study spontaneously produced at the beginning of
the experiment—albeit to the detriment of success,
in that such behavior leads to collisions.
The second qualitative type of movement
trajectories exhibited by Eq. (5) occur when γ
1
≠ γ
2
.
As can be seen from an inspection of the right panel
of Fig. 4, when γ
1
≠ γ
2
an asymmetry in the between
target movement trajectories emerges, as well as a
phase lag between the more elliptical and the more
straight-line trajectory. This behavioral pattern is
similar to the successful task solution adopted by
pairs in the Richardson et al. study and is consistent
with the hypothesis that participants adopted an
asymmetric relation in coupling in order to avoid
collisions and simultaneously synchronize their
between target movements.
Figure 4: Examples of the three qualitative types of
movement trajectories that can be produced by the
Richardson et al., (2015) collision avoidance model, Eq.
(5), for various settings of the parameters of γ
1
and γ
2
(see
text for more details).
Lastly, if γ
1
= γ
2
> 0, then equivalent motion
patterns are created along
and
resulting in
elliptical trajectories that are symmetric across
participants and synchronized with zero phase lag
between the participants’ limit cycle axes.
Interestingly, this third situation also results in a
stable collision avoidance solution, especially for γ
1
= γ
2
>> 0, but one that does not include a phase lag
between the limit-cycle axes and, hence, has a lower
‘margin of safety’ than the asymmetric solution
adopted by participants for the second qualitative
type of movement pattern described above.
The third type of task solution was observed in a
follow-up study (Eiler et al., 2015), in which the
participants were not penalized for collisions. Under
these conditions, participant pairs also produced
movements with less ellipticality. That is, pairs
produced a more symmetrical pattern of elliptical
inphase coordination with a smaller degree of
ellipticality than when collisions were penalized.
Given Eq. (5), this suggests that decreasing the cost
of failure not only weakened the repulsive coupling
between participants, but also resulted in pairs
employing similar magnitudes of repulsive
avoidance (i.e., γ
1
≈ γ
2
> 0).
Most recently, Eiler et al., (2015) have also
demonstrated how Eq. (5) can predict the types of
patterns exhibited between pairs of individuals
walking or running back and forth between target
Modeling Embedded Interpersonal and Multiagent Coordination
159

locations in a real 3-dimensional space. Specifically,
Eiler et al., instructed pairs of participants to walk or
run at a comfortable pace back and forth between
sets of target landmarks positioned in a cross-type
arrangement (see Fig. 5, left). The distance between
the participants’ target landmarks were also
manipulated, with the landmarks positioned either 3
meters or 5 meters apart. Of particular significance,
was that the 2 (speed: walk vs. run) by 2 (distance: 3
vs. 5 m) within subjects design employed in this
study essentially mapped onto different
“chances/severity” of collision conditions, with
walking between targets positioned 5 m apart having
the lowest chance/severity of collision and running
between targets positioned 3 m apart having the
highest chance/severity of collision.
Figure 5: (left) experimental setup for the walking and
running collision avoidance task investigated by Eiler et
al., (2015). (right) Example movement trajectories for the
different target distance and pace conditions explored.
Figure 6: Experimental setup (top) and birds-eye view of
game field (bottom) for the sheepdog game investigated
by Nalepka et al., (2015).
As expected, the degree to which pairs adopted
synchronous straight line or elliptical movement
trajectories between the target locations, as well as
the symmetry of the co-actors movement
trajectories, was a function of these chance/severity
of collision manipulations (see Figure 5, right)—
patterns that could be captured by Eq. (5) by
modulating limit-cycle axis frequency (i.e., k
xi
and
k
yi
) and the magnitude and symmetry of the
repulsive coupling parameters γ
i
.
4 MULTIAGENT HERDING
As a second example of how task dynamic modeling
can be employed to understand EMAD, we consider
the multiagent sheepdog herding game recently
investigated by Nalepka et al., (2015). This game
required pairs of naïve participants to work together
to contain groups of 3, 5, or 7 virtual sheep (i.e.
small, ‘wool’-covered balls) within a central target
region of a virtual field presented on a large tabletop
display (see Fig. 6). The sheep’s’ movements were
governed by random Brownian motion dynamics
and were repelled away from virtual dogs (a blue or
red colored box) that the participants’ controlled in
real-time using hand-held motion tracking sensors.
Specifically, the participants held the sensors in their
hands on top of the colored boxes (dogs) projected
on the tabletop display, essentially making their
hand the “sheepdog”.
Initially all participants pairs adopted a kind of
search and recover strategy, in which each individual
would move toward and corral the furthest sheep from
the center of the game field on their side
However, when a pair’s search and recover
performance improved to the point where they could
consistently and effectively corral the sheep into the
target region in the center of the game field, most
pairs spontaneously transitioned to a coupled
oscillator containment strategy, in which the
participants synchronously moved back and forth in
a semicircular inphase or antiphase manner around
the target containment region—establishing a kind
of “spatiotemporal” wall around the sheep. The two
modes of behavior are illustrated in Figure 7.
Given that nearly all participant pairs
spontaneously adopted the search and recover
strategy at the very beginning of the
experiment(participants could not talk during the
experiment) and then, over the course of repeated
performance, also discovered the coupled oscillatory
containment strategy, Nalepka, et al., concluded that
both behavioral modes were entailed by the
constraints, goals and inter-agent couplings that
defined the multiagent sheep herding game.
COMPLEXIS 2016 - 1st International Conference on Complex Information Systems
160
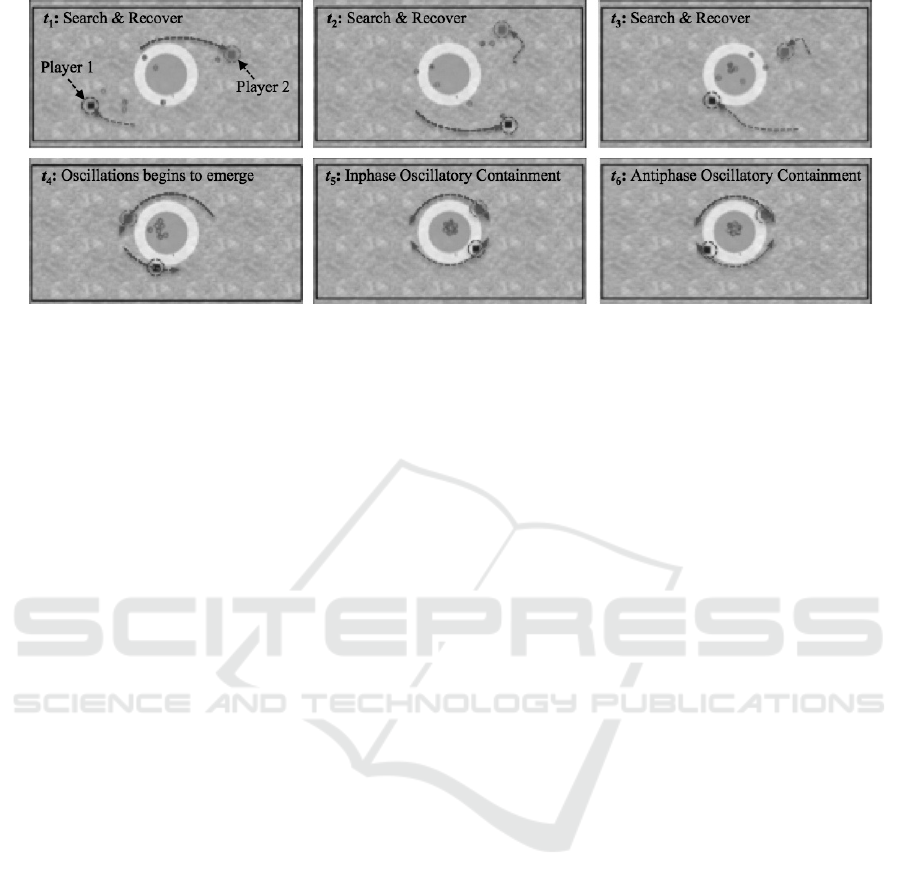
Figure 7: Time lapsed (t1…t6) illustrations of the two behavioral modes exhibited by pairs in the sheepdog herding game.
Dashed arrows indicate movement direction. Dashed circles highlight participant (sheepdog) position of the table, moving
from sheep to sheep in an attempt to get all of the sheep within the central target area and then keep them there.
In other words, the behavioral modes adopted (and
discovered) by participants were both stabilities of
the games task dynamics. Furthermore, finding that
the stable patterns of synchronized oscillatory
containment were consistent with the stabilities of
intra- and interpersonal rhythmic coordination (e.g.,
Haken et al., 1985; Kelso, 1995; Schmidt et al.,
1990; Schmidt et al., 1998; Richardson et al.,
2007)——namely, stable inphase and antiphase
behavior was observed, with inphase coordination
occurring more frequently and more stably than
antiphase coordination—also led Nalepka and
colleagues to conclude that the underlying
dynamical system should entail a corresponding
coupled oscillator process.
In support of these conclusions, Nalepka, et al.
have been developing a simple task dynamic model
of the sheepdog game that not only captures (i) both
behavioral modes within the same system of
equations of motion and (ii) the prototypical
dynamics of interpersonal rhythmic coordination
(i.e., coupled oscillators), but also (iii) the possibility
for a spontaneous transition between the two
behavioral modes as a function of a sheep-distance
dependent Hopf bifurcation (here we present a
preliminary version of the model that has been
developed to date). As illustrated in Fig. 8, the
preliminary model (still to be completely validated)
captures the terminal objective of a participants’
behavior with respect to the center of containment
region within the ξ
x
and ξ
y
game space. Here, x
i
is
the oscillatory perimeter path of each participant i’s
hand, where i = 1 or 2, with respect to half (π-
radians) of the target containment region of success
closest to the participant’s side of the game space. y
i
corresponds to the radial distance of each participant
from the center of the game space and θ
i
is the radial
angle of each participant from the center of the game
space defined with respect to the ξ
y
polar axis on
each participants side of the table (i.e., +ξ
y
for
participant 1 and -ξ
y
for participant 2). Thus, each
participant’s perimeter path, x
i
, is centered on the
participants radial (y
i
, θ
i
) position within the (ξ
x
, ξ
y
)
game space.
To be consistent with the previous research
modeling the dynamics of rhythmic arm and hand
movements (Kay et al., 1987), and rhythmic
interlimb and interpersonal coordination (Haken, et
al., 1985; Schmidt and Richardson, 2008), the
topology of the x
i
perimeter path movement was
defined using a set of coupled hybrid nonlinear
oscillators of the form
+
+
+
+
=
(
−
)
(
−
(
−
)
)
+
+
+
+
=
(
−
)
(
−
(
−
)
)
(6)
with the positive/negative (excitatory/inhibitory)
damping parameters
and
, scaled as a function
of sheep distance using the equation
=
(
,()
−
,
−
)
(7)
For Eq. 6,
and
,
and
,
and
correspond
to the position, velocity and acceleration of each
agent’s hand along the
path,
defines the
stiffness or frequency of movement along the
corresponding
path, and the functions (
) and
(
) corresponding to Rayleigh and van der Pol
escapements terms, respectively. The coupling
function to the right of the equals sign in each
equation is the same as that previously derived by
Haken et al., (1985), and defines both inphase (0°)
and antiphase (180°) relative phase relationships as
the stable coordination modes between the two
Modeling Embedded Interpersonal and Multiagent Coordination
161

oscillators (when
and
< 0), whose relative
strength is defined by the parameters A and B.
Figure 8: Illustration of the task space employed for the
sheep-herding model which captures player i’s (where i =
1 or 2) sheepdog location at any time within the ξ
x
and ξ
y
planar game space in polar coordinates (y
i
, θ
i
), with y
i
corresponding to the radial distance of player i from the
center of the ξ
x
and ξ
y
planar game space and θ
i
corresponding to player i's radial angle (± 90°) from the ξ
y
game space axis on their side of the game field. x
i
is the
oscillatory perimeter path of each participant i’s sheepdog
with respect to half (π-radians) of the target containment
region of success closest to the participant’s side of the
game space, such that each participant’s perimeter path, x
i
,
is centered on the participants radial (y
i
, θ
i
) position within
the (ξ
x
, ξ
y
) game space (see text for more details).
Central to the switch between the search and
recover and oscillatory containment modes of
behavior is the Hopf bifurcation that occurs for each
oscillator system,
, when
is decreased from a
positive to a negative value. That is, when
> 0,
behavior is that of a nonlinear damped mass spring
with a stable fixed-point solution. However, when
< 0,
behavior is that of a nonlinear limit cycle
oscillator.
As defined in Eq. (7), the value of
at any
instance in time, (t), is a differential function of the
distance,
,()
, of the furthest sheep on participant
i’s side of the game space with respect to a
maximum safe containment distance,
,
and a
fixed rate of change parameter
. Accordingly,
when the distance,
,()
, of the sheep furthest
from the center of the game space on participant i's
side of the game space is outside participant i's
maximum safe containment distance,
,
,
> 0
and behavior along the
corresponds to that of a
nonlinear mass damped spring. Conversely, when
the distance,
,()
, of the sheep furthest from the
center of the game space on participant i's side of the
game space is inside participant i's maximum safe
containment distance,
,
,
< 0 and behavior
along the
corresponds to that of a nonlinear limit
cycle oscillator.
A participant’s radial distance,
, was defined as
+
+
−(
,()
+
,
) = 0
(8)
with the radial orientation
defined by
+
+
−
,
(
)
,
=0
(9)
Here,
and
, and
and
correspond to the
velocity and acceleration of participant i’s radial
distance and radial orientation from the center of the
game space, respectively.
,()
is again the
distance of the furthest sheep on participant i’s side
of the game space and
,()
is the angle of the
furthest sheep on participant i’s side of the game
space relative to corresponding ξ
y
polar game space
axis.
,
is a fixed parameter the sets minimum
preferred distance that a participant likes to approach
a sheep, and
and
scale the force (rate) at which
participant i minimizes the difference between the
radial distance and radial angle of their sheepdog
and the radial distance and angle of the furthest
sheep from the center of the game space on their
side of the game field, respectively. Finally,
,
is
a Heaviside parameter defined as
=
0,
,()
,
1,
,()
,
(10)
which results in the stable fixed point solution
,
=0, when all of the sheep on participant i’s
side of the game space are within the participant’s
maximum safe containment distance,
,
. Thus,
when all of the sheep on participant i’s side of the
game space are within the region of containment, the
radial orientation
approaches zero for participant i
and their corresponding
path is centered about ξ
y
.
Collectively, the task dynamic model of the (bi-
agent) sheepherding game can be written as follows,
COMPLEXIS 2016 - 1st International Conference on Complex Information Systems
162

+
+
−(
,()
+
,
) = 0
+
+
−
,
(
)
,
=0
+
+
+
+
=
(
−
)
(
−
(
−
)
)
=
(
,
(
)
−
,
−
)
=
0,
,()
,
1,
,()
,
+
+
−(
,()
+
,
) = 0
+
+
−
,
(
)
,
=0
+
+
+
+
=
(
−
)
(
−
(
−
)
)
=
(
,()
−
,
−
)
=
0,
,()
,
1,
,()
,
(11
)
Importantly, not only does this model effectively
capture the bimodal behavior exhibited by pairs in
the experimental study, but it is also resistant to
perturbations in the sheep movement and location
and is able to spontaneously transition between the
search and recover and oscillatory containment
behavioral modes via a sheep distance dependent
Hopf bifurcation process. Videos presenting
example demonstrations and simulations of the
model, as well as a real participant behavior can be
viewed at: http://www.emadynamics.org/bi-agent-
sheep-herding-game/.
5 CONCLUSION
Our aim here was to provide a brief overview of
how EMAD can be modeled and understood using a
task dynamic framework. It is important to
appreciate that the goal of dynamical modeling is
not to perfectly simulate the exact trajectory or end
state of system behavior, but to shed light on the
structural relations and self-organizing processes
that give rise to effective and robust behavior.
Indeed, the power of a task dynamical model rests
on its ability to validate hypotheses, generate
testable predictions, and motivate future research
questions. It is in this way that developing self-
organized task dynamic models have the potential to
uncover the fundamental processes that shape and
constrain human behavior in general.
ACKNOWLEDGEMENTS
The research was supported by National Institutes of
Health, R01GM105045.
REFERENCES
Chemero, A., 2009. Radical embodied cognitive science.
Boston, MA: MIT Press.
Coey, C., Varlet, M., Richardson, M. J., 2012.
Coordination dynamics in a socially situated nervous
system. Frontiers in human neuroscience. 6, 164.
Dumas, G., de Guzman, G. C., Tognoli, E., Kelso, J. S.,
2014. The human dynamic clamp as a paradigm for
social interaction. Proceedings of the National
Academy of Sciences, 111(35), E3726-E3734.
Eiler, B., Coey, C. A., Ariyabuddhiphongs, K., Kallen, R.
W., Harrison, S. J., Saltzman, E., Schmidt, R. C.,
Richardson, M. J., 2015. Poster presented at the 5th
Joint Action Meeting, Budapest, Hungary, July 2015.
Eiler, B., Kallen, R. W., Harrison, S. J., Saltzman, E.,
Schmidt, R. C., Richardson, M. J., 2015. Behavioral
Dynamics of a Collision Avoidance Task: How
Asymmetry Stabilizes Performance. In Noelle, D. C.,
Dale, R., Warlaumont, A. S., Yoshimi, J., Matlock, T.,
Jennings, C. D., & Maglio, P. P. (Eds.) Proceedings of
the 37th Annual Meeting of the Cognitive Science
Society. Austin, TX: Cognitive Science Society.
Eiler, B., Kallen, R. W., Harrison, S. J., Richardson, M. J.,
2013. Origins of Order in Joint Activity and Social
Behavior. Ecological Psychology, 25, 316–326.
Graf, M., Schütz-Bosbach, S., Prinz, W., 2009. Motor
Involvement in Action and Object Perception
Similarity and Complementarity. In G. Semin, & G.
EchterhoV (Eds), Grounding sociality: Neurons,
minds, and culture. NY: Psychology Press.
Haken, H., Kelso, J. A. S., Bunz, H., 1985. A theoretical
model of phase transitions in human hand movements.
Biological Cybernetics, 51, 347-356.
Kay, B. A., Kelso, J. A., Saltzman, E. L., Schöner, G.
(1987). Space–time behavior of single and bimanual
rhythmical movements: Data and limit cycle model.
Journal of Experimental Psychology: Human
Perception and Performance, 13(2), 178.
Kelso, J. A. S., 1995. Dynamic patterns. Cambridge, MA:
MIT Press.
Knoblich, G., Butterfill, S., Sebanz, N.,
2011. Psychological research on joint action: theory
and data. In B. Ross (Ed.), The Psychology of
Learning and Motivation, 54 (pp. 59-101), Burlington:
Academic Press.
Marsh, K. L., Richardson, M. J., Schmidt, R. C., 2009.
Social connection through joint action and
interpersonal coordination. Topics in Cognitive
Science, 1, 320-339.
Nalepka, P., Riehm, C., Mansour, C. B., Chemero, A.,
Richardson, M. J., 2015. Investigating Strategy
Discovery and Coordination in a Novel Virtual Sheep
Modeling Embedded Interpersonal and Multiagent Coordination
163

Herding Game among Dyads. In Noelle, D. C., Dale,
R., Warlaumont, A. S., Yoshimi, J., Matlock, T.,
Jennings, C. D., Maglio, P. P. (Eds.) Proceedings of
the 37th Annual Meeting of the Cognitive Science
Society. Austin, TX: Cognitive Science Society.
Newman-Norlund RD, Noordzij, ML, Meulenbroek,
R.G.J, Bekkering H., 2007. Exploring the brain basis
of joint action: Co-ordination of actions, goals and
intentions. Social Neuroscience, 2, 48-65.
Richardson, M. J. Dale R., Marsh, K. L., 2014. Complex
Dynamical Systems in Social and Personality
Psychology: Theory, Modeling and Analysis. In H. T.
Reis, and C. M. Judd. (Eds.). Handbook of Research
Methods in Social and Personality Psychology, 2
nd
Edition. New York, NY: Cambridge University Press.
Richardson, M. J., Kallen, R. W., 2015. Symmetry-
Breaking and the Contextual Emergence of Human
Multiagent Coordination and Social Activity. In E.
Dzhafarov, S. Jordan, R. Zhang, and V. Cervantes
(Eds.). Contextuality from Quantum Physics to
Psychology. (pp. 229-286). World Scientific.
Richardson, M. J., Harrison, S. J., Kallen, R. W., Walton,
A., Eiler, B., Schmidt, R. C., 2015. Self-Organized
Complementary Coordination: Dynamics of an
Interpersonal Collision-Avoidance Task. Journal of
Experimental Psychology: Human Perception and
Performance.
Richardson, M. J., Marsh, K. L., Isenhower, R., Goodman,
J., Schmidt, R. C., 2007. Rocking together: Dynamics
of intentional and unintentional interpersonal
coordination. Human Movement Science, 26, 867-891.
Riley, M. A., Richardson, M. J., Shockley, K. Ramenzoni,
V. C., 2011. Interpersonal Synergies. Frontiers in
Psychology, 2, 1-7.
Saltzman, E. L., Kelso, J. A. S. (1987). Skilled actions: A
task dynamic approach. Psychological Review, 94, 84–
106.
Schmidt, R. C., Bienvenu, M., Fitzpatrick, P. A.,
Amazeen, P. G., 1998. A comparison of within- and
between-person coordination: Coordination
breakdowns and coupling strength. Journal of
Experimental Psychology: Human Perception and
Performance, 24, 884-900.
Schmidt, R. C., Carello, C., Turvey, M. T., 1990. Phase
transitions and critical fluctuations in the visual
coordination of rhythmic movements between people.
Journal of Experimental Psychology: Human
Perception and Performance, 16, 227-247.
Schmidt, R. C., Fitzpatrick, P., Caron, R., Mergeche, J.,
2011. Understanding social motor
coordination. Human Movement Science, 30, 834-845.
Schmidt, R. C., O'Brien, B., 1997. Evaluating the
dynamics of unintended interpersonal coordination.
Ecological Psychology, 9, 189-206.
Schmidt, R. C., Richardson, M. J., 2008. Dynamics of
Interpersonal Coordination. In A. Fuchs & V. Jirsa
(Eds.). Coordination: Neural, Behavioral and Social
Dynamics. (pp. 281-308). Heidelberg: Springer-
Verlag.
Schmidt, R. C., Turvey, M. T., 1994. Phase-entrainment
dynamics of visually coupled rhythmic movements.
Biological Cybernetics, 70, 369-376.
Vesper, C., Butterfill, S., Knoblich, G., Sebanz, N.,
2010. A minimal architecture for joint action. Neural
Networks, 23, 998-1003.
Warren, W. H. (2006). The Dynamics of Perception and
Action. Psychological Review, 113, 358–389.
COMPLEXIS 2016 - 1st International Conference on Complex Information Systems
164
