
Social Transmission of Information through Virtual Robotic Agents
Owais Hamid
1 a
, Shruti Chandra
2
, Kerstin Dautenhahn
2,1
and Chrystopher Nehaniv
1,2
1
Systems Design Engineering, University of Waterloo, Waterloo, ON, Canada
2
Electrical and Computer Engineering, University of Waterloo, Waterloo, ON, Canada
Keywords:
Social Learning, Game Environment, Observational Conditioning, Stimulus Enhancement, Response Facili-
tation.
Abstract:
Social learning includes simple or complex social mechanisms that allow us to understand cooperation and
communication in animals, giving them better chances to survive for longer and thrive as a society. Specifi-
cally, certain types of social learning such as observational conditioning and stimulus enhancement have been
investigated in the context of social information spread between primates. However, not many studies have
utilized such social learning mechanisms to study social learning between humans and artificial agents. In the
work described here, we seek to understand if and how simple social learning mechanisms can influence hu-
man participants using an online game platform with an immersive first person experience built through Unity.
Specifically, we designed a study inspired by experiments in behavioural sciences to investigate whether and
to what extent, a robotic agent can influence human’s actions. The study compared two conditions in which
the robot showed body-language based emotions in a positive or negative manner that could enhance certain
stimuli for the human participants and influence their decision making. From this, we wanted to understand
whether these effects are socially learned by humans. Objective (position of player in-game) and Subjec-
tive (questionnaires) measures were recorded, and markers using the objective data suggest successful social
transmission of information. We believe this approach can make a novel contribution to the field of Human
Interaction with Artificial Agents.
1 INTRODUCTION
Social learning is the facilitation of learning by ob-
servation or interaction with another individual or its
products (Hoppitt and Laland, 2013). While these in-
teractions have typically been studied in humans and
other social animals, certain recent literature has fo-
cused on social interaction among generalized agents
that may be artificial, including robots (Steels and Ka-
plan, 2000) (Hamid et al., 2020).
Several mechanisms play a role in the learning
processes that we see every day. Some of them have
been termed cognitively more complex, requiring a
greater amount of cognition such as imitation. Others
are more simple and need not require complex per-
ceptual and cognitive processes, but may involve a
simple redirection of attention, local enhancement be-
ing one good example. In between these two levels of
complexity lie a plethora of social transmission mech-
anisms with varying levels of cognitive complexity
required. Mechanisms that were defined earlier in
a
https://orcid.org/0000-0002-8942-4621
the literature and remain popular include Stimulus-
Stimulus learning which includes Observational Con-
ditioning, the mechanism identified behind Pavlov’s
famous experiments. Emulation is another mecha-
nism that was identified earlier in (Hoppitt and La-
land, 2013). However, while we know that humans
can socially learn from other humans, they may also
learn socially from non-human agents. Humans learn
a variety of skills and behaviours through interaction
with others, cf. language learning through interac-
tion with others (Tomasello, 2009). Social signals
can be non-verbal, or can involve vocal, non-speech
signals. For example, vervet monkeys (Cercopithe-
cus aethiops) use vocal calls for anti-predator alarm
signals, even going so far as to classify which type of
predator is hunting them (Seyfarth et al., 1980). How-
ever, such specialized signalling only works within a
certain species, not across species.
So far, we discussed examples where communi-
cation happens within a species of primates. How-
ever, there do exist several instances of humans ex-
tracting or delivering information socially from or to
Hamid, O., Chandra, S., Dautenhahn, K. and Nehaniv, C.
Social Transmission of Information through Virtual Robotic Agents.
DOI: 10.5220/0010823000003116
In Proceedings of the 14th International Conference on Agents and Artificial Intelligence (ICAART 2022) - Volume 3, pages 361-372
ISBN: 978-989-758-547-0; ISSN: 2184-433X
Copyright
c
2022 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved
361

animals. In (Byrne and Tanner, 2006), this type of
‘imitation’ by an animal (a western lowland gorilla,
Gorilla g. gorilla), who learns to reproduce observed
actions, is considered simple priming where the ani-
mal puts together actions it already knows, and then
reinforces them by individual learning. The gorilla
in this experiment was able to ’imitate’ gestures that
humans showed her to the extent that naive coders
who were asked to check if such actions resembled
the original gestures by humans gave scores that con-
firmed socially learned behaviors. This particular
form of learning, previously called gestural imitation,
was termed in this study as Response Facilitation.
Complex cognitive processes can be involved in
socially learning from others, in particular in the area
of imitation learning. Large bodies of work have been
developed on Imitation and Learning by Demonstra-
tion between robots and humans, where robots are
being taught by human teachers. However, crucial
perceptual and cognitive challenges remain to be re-
solved, and some of them are discussed in (Nehaniv
and Dautenhahn, 2001). This serves as the inspiration
for utilizing simpler methods of social learning so that
robots and artificial agents are able to learn socially in
human-agent interaction.
2 BACKGROUND
A number of efforts have been made to classify so-
cial mechanisms of information transfer in primates,
and the definitions have changed over time. Some of
the older efforts were made by Galef (2013), Heyes
(1994) and Zentall (2001). The ones utilized here are
more recent versions built on previous work (Hoppitt
and Laland, 2013).
Several researchers have studied learning behav-
ior among animals, where a demonstrator directs an
observer’s
1
attention to the specific location of a food
object using simple attention mechanisms such as tap-
ping the item with a body part or partially biting and
chewing a strategically important part that needs to
be exploited initially so that food acquisition by a
‘naive’ observer is made easier (Previde and Poli,
1996; Terkel, 1996). For example, in wild greylag
geese, attention is drawn by tapping a certain loca-
tion. This same effect can be had if a human taps a
certain object in a specific place (Fritz et al., 2000).
1
We use the words demonstrator and observer through-
out the paper to refer to the agent ‘demonstrating’ a certain
behavior in which it is already experienced in, and the naive
agent receiving social information as an ‘observer’ respec-
tively.
In a classic study in humans, Gerull and Rapee
(2002) have documented that affective responses to
novel objects can be learned socially. Specifically, if
humans observe something novel, they usually asso-
ciate certain emotions to the newly introduced object
of interest when they observe someone else who has
more experience react to the novel object or situa-
tion in a certain way. In (Gerull and Rapee, 2002),
toddlers, when first being introduced to novel toy
creatures, learned to associate fear and avoidance be-
havior towards the objects when they witnessed their
mothers reacting to the objects with fear, horror or
disgust and avoidance. Toddlers reacted significantly
differently when a mother’s reaction was positive, not
showing any avoidance to the toy physically. Fa-
cial expressions were also very different in the two
cases, the children in the negative condition showing
expressions of clear disgust or horror, while those in
the positive condition treated the toy normally. This
shows that the information provided by an experi-
enced demonstrator (e.g. mother) is crucial to the per-
ception of any object that the observer (e.g. toddler),
hence creating a Stimulus-Stimulus pairing. Gerull
and Rapee also found significant differences between
the perception and avoidance behavior of male tod-
dlers and female toddlers to the mother expressing a
positive or negative emotion. Male toddlers were less
avoidant towards the toy creature in the negative con-
dition than female toddlers. Furthermore, the impres-
sion of these emotions lasted for at least up to 10 min-
utes, making this one of the first studies to understand
potentially how long the effects might last.
This specific experiment is of great interest to us
for a variety of reasons. New environments pose a
challenging problem when humans who are naive to
the environment first encounter challenges. An expe-
rienced agent that understands the subtleties and prob-
lems of an environment can pass on information to
newcomers, gained by previously exploring this envi-
ronment. We therefore envision situations when em-
bodied artificial agents, quite specifically robots, in-
troduce humans to a new environment efficiently. For
instance, teaching humans about features and situa-
tions in the environment that are dangerous, and oth-
ers that are harmless, even if they may appear danger-
ous. Our study therefore aims to draw inspiration the
above-mentioned social learning study (Gerull and
Rapee, 2002), in the new context of fostering Agent
Interaction and Communication that can be beneficial
to learning about a new environment that the human
encounters. Note that the study by Gerull and Rapee
is itself an extension of Cook and Mineka’s series
of experiments (the original experiments were con-
ducted from 1984-1993, but a comprehensive review
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
362

can be found in (Mineka and Cook, 2013)) which
showed observational conditioning of fear of snakes
among rhesus monkeys, using toy snakes.
2.1 Types of Social Learning
The process of Social Learning is rarely random or
without reason. In fact, social transmission of infor-
mation has been termed strategic. A very complete
list of strategies for social learning can be found in
(Hoppitt and Laland, 2013). Some of the most com-
mon reasons are “when” strategies such as copy when
uncertain. For instance, naive rats copy other more
experienced rats for their choice of diet (Galef, 1996).
Historically, most sub-types of social learning have
focused on imitation, with earlier work considering
all forms of animal based social learning to be an at-
tempt to reproduce human-like imitation (Hoppitt and
Laland, 2013). However, other scientists have catego-
rized the same behaviours as different types of social
learning. Classification schemes for social learning
mechanisms are broadly categorised according to the
following: (1) context specificity (i.e. whether depen-
dent on location or stimulus); (2) whether the out-
come is sensitive to the demonstrator’s actions; (3)
whether the information being transferred is action
specific and; (4) if the learned behaviour is a novel
action sequence (Hoppitt and Laland, 2013, p. 99).
In our study, we are interested in understanding
if social information can be spread by an emotion-
ally intelligent robotic agent to a human, similar to
how mothers’ emotional reactions were transmitted
to their infants (Gerull and Rapee, 2002). In doing
so, we wish to establish a precedent on the effects
that social behavior expressed by non-human artificial
agents can have on humans.
In the kinds of observation based learning that
were discussed above, certain specific types of social
learning stand out and are described and defined as
follows:
Response Facilitation (RF): is defined as the “pres-
ence of a demonstrator performing an act [that] in-
creases the probability of an animal which saw it do
the same” (Byrne, 1994, p. 237). As mentioned in
Section 1, a good example is that when a demonstra-
tor performs such actions, the observers have a “prim-
ing” effect. Note, this effect may be transient, i.e. the
probability of the observers performing the same ac-
tions might reduce later in time.
Observational Conditioning: is one of the older
mechanisms that was discovered through classical de-
signs of social learning studies. The earliest stud-
ies in this category include Pavlovian conditioning
where an Unconditioned Response (UR) to a stim-
ulus for the demonstrator acts as an Unconditioned
Stimulus (US), which then becomes a Conditioned
Stimulus (CS) for the observer, and the observer will
then respond the same way. Heyes’ definition is
a little broader in that stimuli need not be condi-
tioned (Heyes, 1994). It is defined as “a subset of
Stimulus-Stimulus learning where an observer ob-
serving a demonstrator is exposed to a relationship
between stimuli at t1 and the observer’s behavior
changes in a detectable manner at a later time t2”.
Finally, Stimulus Enhancement: as defined by
Heyes (ibid.) occurs “when an observer observing
a stimulus at time t1 from a demonstrator and this
stimulus causes an observable change in the observer
at time t2”. Stimulus enhancement requires memory
and learning, since the observer has to retain informa-
tion about the object and how it was used, in order to
use the same object in a different context, or a differ-
ent object in a similar context(Giraldeau, 1997).
It should be noted that there exists plenty of
overlap between the above-mentioned types of social
transmission mechanisms. This is due to the reason
that a combination of factors might be at play when
we see certain types of imitation-like behaviors in an-
imals.
2.2 Social Learning with Artificial
Agents
Little research has been conducted on how artificial
agents interacting with humans might shape or in-
fluence their experiences. This is specifically the
case for non-verbal social information spread not in-
cluding cognitively complex forms of imitation. A
certain amount of literature review on social learn-
ing in robotics, specifically multi-agent robotics was
conducted in (Hamid et al., 2020). Cakmak et al.’s
work is one of the few examples that can be found
of human-robot interaction with social mechanisms
simpler than imitation (Cakmak et al., 2010). It in-
vestigates how different social learning mechanisms
such as emulation, mimicking and stimulus enhance-
ment (three other simpler forms of social transmis-
sion) can be used to teach robots certain simple tasks.
Similarly, social learning was also utilized for ba-
sic language learning using the Aibo robot (Cakmak
et al., 2010). While no specific type of social trans-
mission mechanism was identified, social learning for
language acquisition through a human mediator is re-
alized through observational learning. Additionally,
supervised and unsupervised machine learning algo-
rithms were used for this task.
Social Transmission of Information through Virtual Robotic Agents
363

Figure 1: A decision tree describing which types of social transmission are at play depending on the effect of the stimulus and
the demonstrator’s actions, adapted from (Hoppitt and Laland, 2013). The social learning mechanisms are classified according
to different levels: if the effect of the stimulus is Context Specific, if it is dependent on the demonstrator’s actions, if the action
itself plays a large role in the effect, and if the effect depends on an action sequence that is novel. PI = Product Imitation, CI
= Contextual Imitation, ORSL = Observational R-S Learning, OC = Observational Conditioning, LE = Local Enhancement,
RF = Response Facilitation, SE = Stimulus Enhancement.
In our work, we investigated simple forms of so-
cial learning mechanisms that could be transmitted by
the artificial agents such as robots to humans. Addi-
tionally, we wanted to understand if robots are just as
effective as experienced demonstrators in conveying
social information to humans.
2.3 Using Virtual Environments as a
Stand-in for Real World
Experiments
Since the platform used for conducting this experi-
ment is entirely virtual, using a game environment, it
is worth asking whether this study can be emulated in
real world conditions, and how close the study gets
to evoking actual emotional responses from partici-
pants. The pertinent practical question being, can we
compare emotional arousal in games to experiencing
emotions in the real world?
While there are different opinions about the extent
of the human reaction to events in digital media (in-
cluding games and movies), (Reeves and Nass, 1996)
pointed out that the responses to media events are re-
markably similar to real world responses. While there
is some agreement that the immediate emotional re-
sponse is very similar, the longer term effects might
be more blunted for virtual events. (Cantor, 2009) as-
serts that previous experiences do play a strong role
in evoking emotional responses, especially from the
real world, however in the virtual world these are less
intense than the same happening in the real world.
(Cantor, 2009) and (Lynch and Martins, 2015) argue
that previous experiences may allow participants to
cope with fear based scenarios more effectively and
allow a more balanced reaction towards these events.
Given the current trend of artificial agents play-
ing increasingly bigger roles in human society, under-
standing how these agents may influence humans is an
important question to consider. We address this ques-
tion by designing a virtual scenario where a robotic
agent needs to guide a human who is new to the en-
vironment, assuming that these might be some of the
roles given to intelligent agents in the future.
3 RESEARCH QUESTIONS &
HYPOTHESES
Despite drawing inspiration from the above men-
tioned experiment that involves a mother transmitting
social information to a child about a novel stimulus
in the environment (Gerull and Rapee, 2002), several
factors have been changed to better suite the study
currently conducted. First, we focus on adults rather
than infants because our use case, i.e. familiarizing
individuals to new environments, suites adults. In the
case of grown adults, they do not necessarily react to
surprising stimuli in a new environment in a manner
similar to how toddlers or infants react. For example,
adults may have inherent biases from previous experi-
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
364

ences that might make them more or less susceptible
to either positive or negative actions. Second, adults
are already used to a large variety of different objects
and experiences, so it can be difficult to create truly
novel stimuli that they previously have never encoun-
tered, making our task of replicating the experiment
difficult. To counter this, we attempted to make the
animal novel and thus chose an alien animal that is
not common in general, does not exist in reality, and
resembles a combination of two or three animals. Ta-
ble 1 outlines the similarities and differences in our
approach as compared to (Gerull and Rapee, 2002).
In this study we are interested in understanding if
social information (in this case affective behaviour by
a virtual robot) can spread to humans who share the
environment (designed similar to a computer game)
with the virtual robot. We study the role of simple so-
cial transmission mechanisms such as Observational
Conditioning, Stimulus Enhancement and Response
Facilitation. Figure 1 portrays the decision making
process that allows us to narrow down these mecha-
nisms.
The methodology of the experiment is described
in detail in Section 4.1, however a functional descrip-
tion of the experiment is given here. The experimen-
tal design requires a novel stimulus in an environment
unfamiliar to the participants, with a virtual robotic
agent acting as a demonstrator. The game was de-
signed such that a novel stimulus would startle the
demonstrator (robotic agent) in a programmed fash-
ion, with the observer constrained to watch the ex-
change in a virtual forest environment. The stimu-
lus chosen had to be novel to some extent, therefore
an “Alien Animal” was chosen for this task. The
demonstrator reacts either positively
2
or negatively
3
to the animal, i.e. acting in either a friendly manner
or running away in terror, respectively. All the while,
the participant/observer watches this reaction of the
demonstrator towards the novel stimulus without be-
ing able to move. The study used a between-subjects
design, i.e. participants saw either the demonstrator’s
positive or negative reaction.
In this context, and with reference to the decision
tree shown in Figure 1, the effect is stimulus specific,
sensitive to the demonstrator’s actions, action spe-
cific, but not a novel action sequence
4
. Thus, we are
2
The actions taken by the virtual robotic agent for the
positive condition can be seen by clicking HERE
3
The actions taken by the virtual robotic agent for the
negative condition can be seen by clicking HERE . It is to be
noted that the alien animal’s actions were exactly the same
in both conditions. Only the demonstrator’s actions were
changed. Further, care was taken to show the same actions
by the animal in both conditions
4
We assume that adult participants are fully capable of
left with three possibilities to classify the social trans-
mission that might occur in this study, namely, ORSL
(Observational Response-Reinforcer Learning), RF
(Response Facilitation) + OC (Observational Condi-
tioning) or RF + SE (Stimulus Enhancement) which
is the red box, second from left in Figure 1. In or-
der to differentiate between these 4 types of social
transmission mechanisms, two factors are important;
whether the observer observes the demonstrator get
rewarded for its behavior, and whether this learn-
ing is S-S (Stimulus- Stimulus) or R-S (Response-
Reinforcer). From our earlier discussion
5
in Section
2, SE, OC and RF do not typically involve rewards
6
.
ORSL can therefore be ruled out. This narrows down
the possible social transmission mechanisms to either
RF + OC or RF + SE. Further information is provided
once the results are explained in Section 6.
We designed a between-subject study that aims to
investigate the effect of a robotic demonstrator on an
observer
7
in an unknown terrain, i.e. a virtual forest.
These characteristics include two affective behaviours
(positive and negative emotions expressed through
body movements) portrayed by a robotic agent when
encountering a novel creature. Note, in order to pro-
vide context for the demonstrator agent’s skills, it is
introduced as having prior knowledge of the environ-
ment, which is new to the observer. We formulated
our research questions as follows:
• RQ1: Does the perceived reaction (positive, neg-
ative) of the demonstrator (robot) affect the ob-
server’s (human participant’s) response? If so, to
what extent?
• RQ2: Does the participants’ gaming experience
or gender affect their responses towards the crea-
ture? If so, to what extent?
With respect to these research questions, we pro-
pose the following hypotheses:
• H1 for RQ1: Distance of the participant to the an-
imal in the positive condition should be lower than
in the negative condition. We expect participants
in the positive condition to have a more positive
attitude towards the alien animal. In (Gerull and
Rapee, 2002), children observe and infer the na-
ture of the relationship between their carer and the
recognizing the demonstrator’s actions. Hence, this effect
is not based on a novel action sequence.
5
A detailed version of these subtleties is discussed
in (Hoppitt and Laland, 2013), Section 4.2
6
There is some debate regarding usage of reward for
OC, see (Palameta and Lefebvre, 1985). However, OC com-
bined with RF may not necessarily involve rewards
7
We use the word observer and participant interchange-
ably since in this experiment, the observing agents are the
human study participants.
Social Transmission of Information through Virtual Robotic Agents
365

Table 1: Description of similarities and differences of the present study, as compared to (Gerull and Rapee, 2002).
Gerull and Rapee Current Study
Mean Age 17 months 30.6 years
Type of Social Transmission Observational Learning Observational Conditioning
Number of Participants
30 44
Gender of Observers
F-15, M-15 F-17, M-27
Demonstrator Mother (Human) Robotic Agent
Stimulus Snake/Spider toy Virtual Alien Animal
Measurement Subjective - Likert Scale Subjective and Objective
Number of times observer
is exposed to stimulus
3 times
1st at 1 min,
2nd at 2 min,
3rd at 10 min
2 times
1st at 1 min,
2nd at 7 min
novel creature by recognizing disgust or fear, and
replicating the same avoidance behavior towards
these creatures.
• H2 for RQ1: Participants in the positive condi-
tion should have a more positive and less violent
perception of the creature as compared to the neg-
ative condition.
• We do not present any hypotheses regarding RQ2
since the question for the effect of gender was
only included because Gerull and Rapee found an
effect, although this was for toddlers. We include
the effects of gaming in the RQ since we would
like to know if previous experience has a signifi-
cantly different effect on a participant’s behavior
in-game.
4 METHODOLOGY
We used Amazon Mechanical Turk to carry out this
remote study. The virtual environment is set in an
unknown terrain (a forest). The game scene includes
a virtual robotic agent capable of two types of emo-
tional body expressions (positive, negative) portrayed
by the robotic-like agent when subjected to a novel
creature, with the robotic agent being depicted as
someone with experience in the environment through
the game narrative. For the participants, the goal of
the game is to collect spheres scattered around the
game world. The spheres were distributed around the
environment in such a way that it would not bias par-
ticipants towards either moving towards or away from
the alien animal. The entire participant experience
was divided into three parts:
• The pre-game Questionnaires: This includes
some basic demographic information, consent and
general information regarding the game. This was
administered using Qualtrics
©
(2021 Qualtrics).
• The game experience included three levels:
– Level 1: Familiarization phase where partici-
pants get used to the game interface using their
keyboard and mouse on a PC/Laptop
– Level 2: Participants explore the area, look-
ing for in-game rewards. The event with social
transmission where the robot reacts either neg-
atively or positively to the alien animal happens
then. Next, the animal is removed from the
game (in accordance with the methodology fol-
lowed in (Gerull and Rapee, 2002)). This level
then continues for another 6 minutes, to make
sure there is no immediate emotional carry-over
to the next phase.
– Level 3: The participant starts the game at a
position where they observe the alien animal
directly. The creature remains at the same po-
sition and does not move in space, it is sim-
ply looking around. The participants can now
explore the area in the vicinity of the animal,
as well as moving further away, picking up re-
sources. This level lasts 7 minutes before the
game is bought to a close.
• The post-game Questionnaires: A set of custom-
made questions ask about participants’ experi-
ences during the game, notably their perception
of the alien animal, and including a few questions
as attention-checks. This questionnaire was also
hosted on Qualtrics.
This study was approved by the Office of Research
Ethics at University of Waterloo.
4.1 Game Development
The game was designed with the Unity 2019 editor.
Participants were asked to enter their ID, and their
in-game positions were recorded once every second.
The game is hosted through the Web Graphics Library
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
366

(WebGL) format on Github pages. This allows the
game to be hosted remotely and to be playable on a
web browser without the user having to install any
plugins, and multiple participants can play simultane-
ously. In-game data is stored on an external online
DataBase based on a MongoDB server on the cloud.
4.2 Questionnaires
Two standardized and two custom-made question-
naires were included in the study and administered
through the University of Waterloo’s Qualtrics sys-
tem, along with the accompanying consent form at
the beginning, an explanation of the tasks, instruc-
tions to play the game and a web page explaining the
exact purpose of the game at the end of the experi-
ment. Custom-made pre-game and post-game ques-
tionnaires were completed by participants. Attention
check questions were used to address some of the is-
sues regarding Amazon Mechanical Turk studies such
as lack of attention or misrepresentation (MacInnis
et al., 2020) (e.g. Have you encountered the creature?
How does the creature look like?). Other questions
concerned participants’ reaction to and perception to-
wards the alien animal in relation to what they ob-
served from the virtual robot demonstrator (e.g. How
did you perceive the behaviour of the robot?).
4.3 Participants
A total of 49 successful participants were recruited
on Amazon Mechanical Turk. Data integrity was im-
portant since there was some packet data loss through
the three-step process with respect to the participants’
position. In order to enhance the quality of the data
collection, participants were recruited with respect to
specific metrics (>96% completion rate for at least 50
previous tasks completed).
The participants were equally distributed accord-
ing to the two experimental conditions. However, due
to some participants failing the attention checks, the
study finally had 27 participants in the negative con-
dition and 22 in the positive condition. Of the 27 in
the negative condition, 3 were excluded due to packet
data loss regarding their location in-game. A further
2 were excluded due to low quality of submissions.
A low quality submission was defined as the partici-
pant either standing at the same place for a long time
(over 3 minutes of the total 6 minutes in the first level,
hence showing inactivity), or answering the question-
naire by clicking on the same options for multiple
questions. This left us with 22 participants for each
condition, in total 44 participants, 28 males and 16
females, 19 to 55 years of age, with the mean age
being 30.64, S.D. of 8.9 years, 32 gamers and 12 non-
gamers.
4.4 Statistical Analyses
To understand the effect of social learning mecha-
nisms on the participants, we collected the data in the
form of objective and subjective measures. The ob-
jective data collected during the study in the second
phase includes mean and absolute distances
8
from the
participant to the alien creature, the frequency of re-
turns of the participant (moving back towards) to the
creature, and the time duration that the participants
took to their first return to the creature. Since there is
no way to directly measure (objectively) how a partic-
ipant feels regarding the animal, we believe distance
of the participant to the animal in-game represents a
faithful marker regarding how wary they feel towards
the animal. A ‘return’ is defined as the participant
walking to a position where they can clearly observe
the animal from a distance of 40 game distance units.
40 is chosen as the cut-off since the landscape con-
sists of hills near the position of the creature where
a participant might be behind one of them and there-
fore not directly have an animal in their line of sight
despite close proximity. These distances are calcu-
lated for each participant in relation to the creature at
different time points: 15 sec; 30 sec; 45 sec; 60 sec
and 100 sec. Because the alien animal was not a real
threat to the participants’ in-game character and al-
ways stood in one place, participants became curious
about the animal after about 100s and started wan-
dering closer to the animal. Therefore, the important
period was the first 100s of the participants observing
the animal and deciding whether to move away or ex-
plore around the animal. We explored different time
stamps from 15 to 100, but from 60s up until 100s,
no difference in results is found. Table 2 describes
the normality tests that were performed to determine
whether a non-parametric or parametric test was re-
quired to test for significance differences between the
positive and negative conditions. Shapiro-Wilk tests
were performed to understand normality of the objec-
tive data. This test checks whether a distribution is
non-normal, i.e. if p<0.05, the data is not normal. If
the data is indeed non-normal, the Mann-Whitney U
test was performed, and the independent-samples t-
test on the data that were normally distributed.
The subjective data includes a question regard-
ing the participant’s perception of the behaviour of
8
For mean distance, we averaged the distance of a par-
ticipant to the animal up to a certain time interval, for ex-
ample, 30 seconds. The absolute distance is the distance to
the animal at that point in time, for instance, at 30 seconds.
Social Transmission of Information through Virtual Robotic Agents
367

(a)
(b)
(c)
Figure 2: a A screenshot of the game showing the robot introducing itself to the participant. A game world map in a circle can
be seen on the top right. bThe Alien Animal that was present in the game world. c An overview of the world. The dirt paths
can be observed crisscrossing the landscape, while towards the bottom left, an overhead view of the alien animal can be seen.
the creature. The question that the participants were
asked was: How did you perceive the behaviour of
the creature?. Two example responses were: ”I per-
ceived the behaviour as curiosity” and ”It seemed vi-
olent”. The responses of all the participants were
classified into three categories of perceived threat:
Mild, High and None. Two independent coders not
involved in the study rated the responses of the par-
ticipants according to the three categories. Cohen’s κ
was run to determine if there was agreement between
the rater’s judgement on whether the participants per-
ceived high, mild or no threat from the alien animal.
There was good agreement between the two judge-
ments, κ = .726 (95% CI, .556 to .896), p ¡ .001.
One other pertinent question asked in the post-
game questionnaire was about the participants’ reac-
tions when they first saw the creature. This is in ref-
erence to Level 2 (see Section 4) when the partici-
pants first saw the creature and the robot’s reaction to
it. The responses were one of [Run Away, Approach,
Neither, Other] and this data was classified as Nomi-
nal, and therefore the Chi-Square test was used with
z-tests for post-hoc analysis.
5 RESULTS
Mean Distance between Participants and Alien
Animal: A Mann-Whitney U test was run to deter-
mine if there were differences in mean distance score
between positive and negative conditions. Mean dis-
tance score at time point 30 sec, was statistically sig-
nificantly higher in the negative condition (Mdn =
57.58) than in the positive condition (Mdn = 48.40),
U = 152, z = -2.113, p = .035, using an exact sam-
pling distribution for U (2-tailed). Similarly, at time
points (for Mean distance) 45 sec and 60 sec, a Welch
t-test was run to determine if there were differences in
distance score between the positive and negative con-
dition. The mean distance for the participants from
the creature in the negative condition was higher (at
45 sec: M = 76.07, SD = 36.11; at 60 sec: M = 88.91,
SD = 42.43 ) than the participants in the positive con-
dition (at 45 sec: M = 54.49, SD = 19.00; at 60 sec:
M = 62.64, SD = 21.12), a statistically significant dif-
ference was found for both time points 45 sec, M =
21.58, 95% CI [-39.31, -3.85], t(31.79) = -2.481, p =
.019, d= -.52 (Moderate effect) and 60 sec M = 26.27,
95% CI [-46.89, -5.65], t(30.80) = -2.6, p = .014, d=
-.55 (Moderate effect).
Absolute Distance between Participants and Ani-
mal: A Mann-Whitney U test was run to determine if
there were differences in absolute distance score be-
tween positive and negative conditions. The absolute
distance scores at time point 15 sec were statistically
significantly higher in the negative condition (Mdn =
56.36) than in the positive condition (Mdn = 46.75),
U = 153.50, z = -2.078, p = .037, using an exact
sampling distribution for U (2-tailed). The absolute
distance scores at time point 30 sec were statistically
significantly higher in the negative condition (Mdn =
70.08) than in the positive condition (Mdn = 51.06),
U = 175=4.00, z = -1.596, p = .013, using an exact
sampling distribution for U (2-tailed).
Perceptions of Participants towards the Behavior
of the Animal: With respect to the first impression of
participants towards the creature, a chi-square test of
homogeneity was run, with an adequate sample size
established according to Cochran (Cochran, 1954).
The two multinomial probability distributions were
equal in the population, χ
2
(3) = 8.581, p = .035. Par-
ticipants in the negative condition were more likely
to respond with “Run Away” (n = 19, 86.4% versus
n = 12, 54.5%). Post hoc analysis involved pairwise
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
368

Table 2: Mean and Absolute distances of participants to the animal at 15, 30, 45, 60 and 100s. For Absolute distance at 60s,
where the positive was normal and the negative non-normal, the Mann-Whitney was used, since both distributions need to be
normal for the T-test. Here N- refers to Non-Normal, and N+ to Normal.
15s 30s 45s 60s 100s
Mean
Positive p<0.05, N- p<0.05, N- p>0.05, N+ p>0.05, N+ p>0.05, N+
Negative p<0.05, N- p<0.05, N- p>0.05, N+ p>0.05, N+ p>0.05, N+
Absolute
Positive p<0.05, N- p<0.05, N- p>0.05, N+ p>0.05, N+ p>0.05, N+
Negative p<0.05, N- p<0.05, N- p>0.05, N+ p<0.05, N- p>0.05, N+
(a) (b)
Figure 3: (a) The mean distance of participants to the alien animal up until 30s, 45s and 60s, respectively, showed a significant
difference between positive and negative conditions. (b) The absolute distance of participants to the animal up until 15s and
30s showed a significant difference between positive and negative conditions.
comparisons using multiple z-tests of two proportions
with a Bonferroni correction. Statistical significance
was accepted at p < .0125. With the Bonferroni Cor-
rections, there were no statistically significant differ-
ences in any of the cases. While the ‘Run Away’ re-
sponse for Positive vs Negative Condition is statis-
tically significant without the Bonferroni correction
(χ
2
(1)=.021), with the correction this is still greater
than .0125.
A Mann-Whitney U test was run to determine if
there were differences in subjective rating scores be-
tween positive and negative conditions for the ques-
tion “How did you perceive the behaviour of the crea-
ture?”. No significant results were found for either of
the raters’ scores between Positive and Negative con-
ditions.
Gaming Experience: Mann-Whitney U tests were
run to determine if there were differences in the dis-
tances between gamers in the positive and negative
conditions. No statistically significant differences
were found for either Mean or Absolute distances.
This was extended to Mann Whitney U-tests between
gamers and non-gamers without regard for the con-
dition they came from. No significant results were
found for this test either.
A Mann-Whitney U test was also run to determine
if there were differences in subjective ratings score
between Gamers in the two conditions. No signifi-
cant results were found for either of the raters’ scores
between the two conditions. Further, subjective rat-
ing scores between gamers and non-gamers was also
non-significant.
Effect of Gender: The Mann-Whitney U tests were
run to determine if there were differences in the dis-
tances according to gender. None of these data points
for either Mean or Absolute distance were found to be
statistically significantly.
Further, the Mann-Whitney U test to determine if
there were differences in subjective rating scores be-
tween the self-reported Male and Female genders (no
other genders were reported) found no significant re-
sults for either of the raters’ scores between Male and
Female participants.
Frequency of Return to Animal: An analysis was
conducted to understand whether participants en-
gaged in searching the area around the animal for re-
wards and returning to it. As before, the definition of
‘return’ was given as approach of participant to within
40 distance units, i.e. within observable distance.
A Mann-Whitney U test was run to determine if
there were differences in the number of times partici-
pants return to the animal in the Positive and Negative
Conditions. The statistic for frequency of returns was
statistically significantly higher in the negative condi-
tion (Mdn = 1, Mean Rank=25.73) than in the positive
condition (Mdn = 2, Mean Rank=19.27), U = 171, z =
-1.722, p = .045, using an exact sampling distribution
for U (1-tailed).
Social Transmission of Information through Virtual Robotic Agents
369

6 ANALYSIS & DISCUSSION
It is essential to interpret the results detailed in Sec-
tion 5 within the context of the definitions provided
in Section 2 and understand if the results support our
stated hypotheses within the definitions laid out ear-
lier.
Objective Measures for Positive & Negative Con-
ditions: A closer look at the histograms in Fig-
ure 4 provides an interesting trend where we observe
the distance of participants in positive condition (blue
bars) to be less spread out and closer to the animal
(higher towards the left side of the histograms), which
represents a lower distance to the alien animal. This
is visible more clearly for 30s, 45s and 60s in Row
1. The differences in the mean distances can be seen
more clearly in Figure 3a that shows significant dif-
ferences between the two conditions.
A similar trend emerges when we compare abso-
lute distances between the two conditions. Histogram
bars in Row 2 of Figure 4 show that participants in
the positive condition are in higher numbers at shorter
distances from the animal, i.e. there are more partic-
ipants closer to the animal in the positive condition
for at least 15s and 30s. This trend disappears as time
stretches further. This means they tend to be closer to
the animal during the earlier stages than the partici-
pants in the negative condition. We hypothesize that
this is because participants start exploring the area
more randomly in search of rewards as time goes on.
The trend of significant differences between partici-
pants in the two conditions is shown in Figure 3b.
One other point of interest is the result from the
frequency of number of returns, as presented in Sec-
tion 4.4. This frequency was calculated to provide an
understanding of how comfortable participants felt in
searching for rewards closer to the alien animal. As-
suming participants from the positive condition return
to the animal more often due to having been exposed
to a prior positive relationship, the 1-tailed test is sig-
nificant. This confirms Hypothesis 1 for Research
Question 1, i.e. the distance of the participant to the
animal in general is significantly lower in the positive
condition than the negative.
Subjective Measures: The subjective data that was
collected with respect to independent coders’ ratings
of the threat level that participants expressed towards
the animal was non-significant. While the results
suggest that there is an effect of the demonstrator’s
(robot) reaction on the participant, these results are
not significant. We hypothesize that this is due to hes-
itance among participants to admit fear in general, and
they may not agree that they felt frightened even if
their in-game behavior shows this. This means that
Hypothesis 2 for Research Question 1 cannot be ver-
ified. We consider subjective data to be supporting
data, and secondary to the objective data.
Gender and Gaming Experience: A close look at
the relevant Sections regarding previous gaming ex-
perience and effect of Gender in Section 4.4 show
us that neither gender nor prior gaming experience
made a significant difference in participants’ attitudes
towards the alien animal. Since no differences can
be found between gamers and non-gamers for either
the subjective or objective data, our data suggests that
gamers and non-gamers have a similar experience in
our study. Furthermore, gamers in the positive and
negative conditions also do not seem to have had a
very different experience. This is suggested by nei-
ther the objective nor subjective data for the positive
or negative conditions being significantly different.
General: Taking the three types of objective data
measures together suggests that participants in the
positive condition tend to stay closer to the animal,
and the only difference between the games played by
participants in the two conditions is the type of reac-
tion to the demonstrator.
With this in mind, referring back to the litera-
ture in social transmission among humans and other
animals, Heyes’ definition of Stimulus Enhancement
(Heyes, 1994), where an observer observing a stimu-
lus at time t1 causes an observable change in the ob-
server’s behavior at time t2, is satisfied. Further, it
can be argued that the observer participants are ex-
posed to a stimulus relationship (fear or positive re-
actions towards a stimulus, i.e. an alien animal) at t1,
which then leads to a similar manner being adopted
by the observers at t2, thus fitting the definition of
Observational Conditioning as well. To provide sup-
port that Response Facilitation has happened, we have
to show that the probability of an observer doing the
exact same thing that it saw the demonstrator doing
must increase. Two problems can arise while trying
to conform to this definition. First, we cannot cal-
culate the probability of adoption of the same action
because we must first define what the ‘same’ action
is. Second, we must demonstrate such a probability
increases, which we cannot do since the repertoire of
actions that the participant can take through the game
is very limited. Hence, it is difficult to provide sup-
port for RF through our online experiment, and so this
theoretical issue remains inconclusive and has to be
investigated in future research.
Of particular importance to theories of fear ac-
quisition, negative stimuli pairing created a negative
impression that lasted over 6 minutes, which was the
time given between Level 2 and Level 3. This means
that the effect lasts at least 6 minutes and does not
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
370
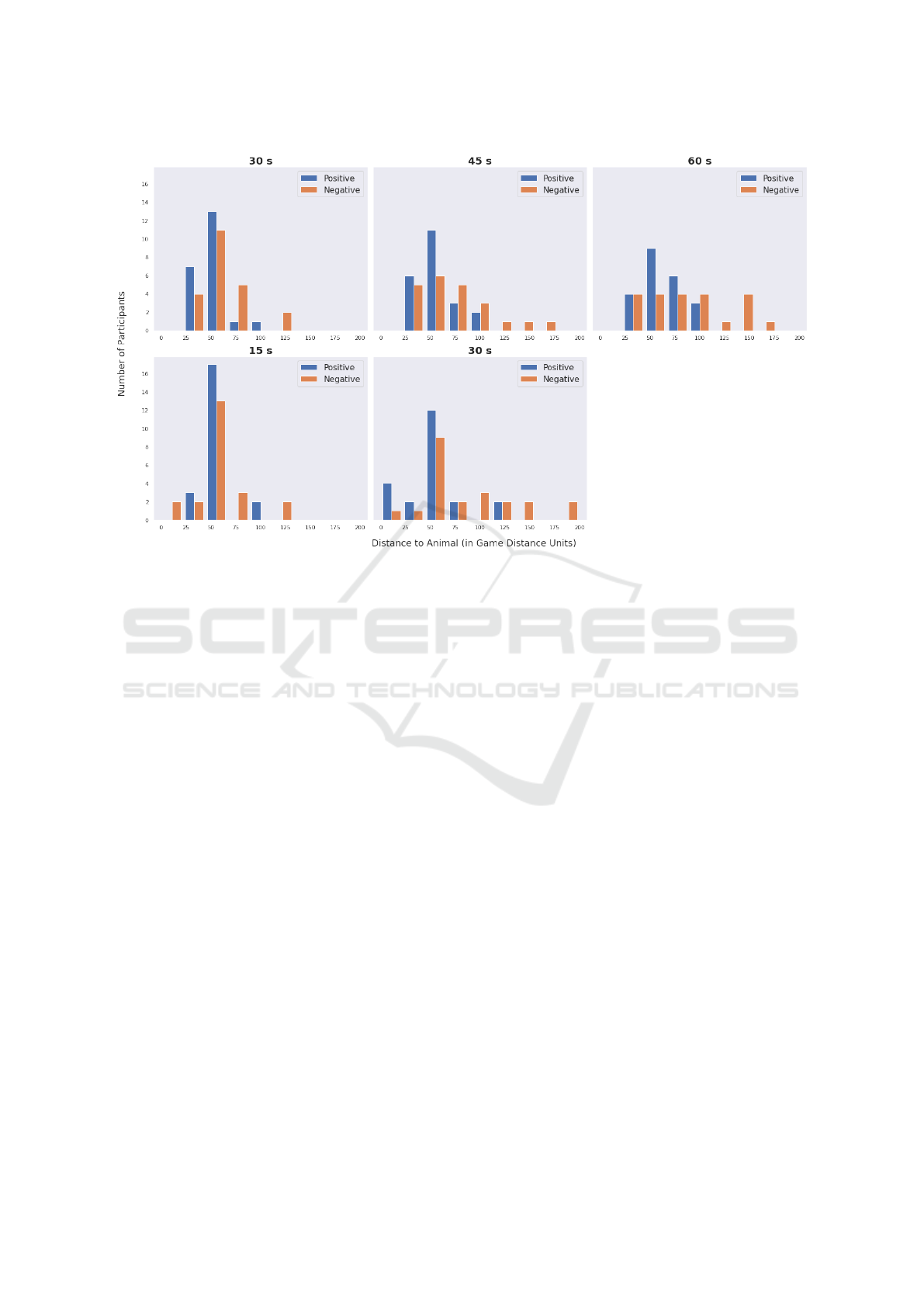
Figure 4: Row 1: The mean distance of participants to the alien animal up until 30s, 45s and 60s, respectively, showed a
significant difference between positive and negative conditions. Row 2: The absolute distance of participants to the animal up
until 15s and 30s showed a significant difference between positive and negative conditions.
simply disappear after the initial appearance of the
alien animal.
As a comparison to the earlier study by (Gerull
and Rapee, 2002), this study suggests the effect of
Observational Conditioning and Stimulus Enhance-
ment to last at least 6 minutes instead of 10 minutes
in the earlier paper. The sample size for this study
includes adults instead of children, is more numerous
(44 versus 30), and is virtual in nature with an artifi-
cial agent as a demonstrator.
7 LIMITATIONS AND FUTURE
WORK
The study is limited in nature for two reasons. First,
it is very difficult to find other studies in human-
human social transmission that can be replicated for
Human Agent Interaction where the agent is artifi-
cial. Therefore, the scope is limited to suggesting
that such types of social transmission are detectable
and quantifiable. Second, while original plans to con-
duct this study were in person, the complete halt of in-
person activities during COVID-19 forced this study
to be conducted online. We would like to note that
as is presented in Section 2.3, since virtual media and
real world experiences are to some extent comparable,
there is a good likelihood that the results produced in
this study can be reproduced using real world scenar-
ios. Therefore, these two limitations become scope
for future work, i.e. the goal would be to conduct in-
person experiments with real robots for human-robot
interaction in order to verify the results gained in this
remote study, and also to expand the studies to further
incorporate other types of social transmission mecha-
nisms. Certain other limitations in this study include
unequal sample sizes regarding gender and gaming
experience, which again was due to limitations of us-
ing crowd-sourcing methods, however, taken care of
by the chosen statistical tests.
While two experimental conditions were studied
here, it would also be desirable to compare these two
conditions with a neutral condition. While we drew
inspiration from (Gerull and Rapee, 2002) who com-
pared only the positive and negative conditions, others
(Mineka and Cook, 2013) utilize the approach with
a neutral condition as well. Finally, variations of
the virtual environment, tasks, and the nature of the
demonstrator (e.g. whether a robotic or human-like
or animal-like agent) could be studied further. All of
these factors make for exciting possibilities to con-
duct future work.
Social Transmission of Information through Virtual Robotic Agents
371

8 CONCLUSION
An online study was designed to understand if sim-
pler forms of social learning (i.e. simpler than imita-
tion) can be observed between artificial agents and hu-
man participants being present in a virtual, game-like
experimental environment. Results from objective
and subjective data collected during the online game,
which was carefully designed to perform stimulus
pairing, point towards successful social transmission
between a robotic virtual agent and human partici-
pants of information utilizing a mixture of methods,
as identified in the literature as Observational Con-
ditioning, Stimulus Enhancement and possibly Re-
sponse Facilitation. The study closely emulates work
done previously in the form of mother-child interac-
tion, (Gerull and Rapee, 2002), and to some extent,
human-animal interaction (Mineka and Cook, 2013).
Neither gender nor previous gaming experience seem
to play any significant role in the efficacy of social
transmission of information in our study.
The study, to the best of our knowledge, is a novel
approach in the field of Human Interaction with Arti-
ficial Agents, inspired by experiments in behavioural
sciences. Further studies with in-person participation
and real robots would be beneficial, once such re-
search is possible, to verify and extend the results.
REFERENCES
Byrne, R. W. (1994). The evolution of intelligence. In
Behavior and Evolution, pages 223–265. Cambridge
University Press, New York, NY, US.
Byrne, R. W. and Tanner, J. E. (2006). Gestural imitation
by a gorilla: evidence and nature of the capacity. In-
ternational Journal of Psychology and Psychological
Therapy, 6(2):215–231.
Cakmak, M., DePalma, N., Arriaga, R. I., and Thomaz,
A. L. (2010). Exploiting social partners in robot learn-
ing. Autonomous Robots, 29(3):309–329.
Cantor, J. (2009). Fright reactions to mass media. In Media
Effects, pages 303–319. Routledge.
Cochran, W. G. (1954). Some methods for strengthening
the common χ
2
tests. Biometrics, 10(4):417–451.
Fritz, J., Bisenberger, A., and Kotrschal, K. (2000). Stim-
ulus enhancement in greylag geese: socially medi-
ated learning of an operant task. Animal Behaviour,
59(6):1119–1125.
Galef, Jr., B. G. (1996). Social enhancement of food prefer-
ences in norway rats: a brief review. In Heyes, C. M.
and B. G. Galef, J., editors, Social Learning in Ani-
mals: The Roots of Culture, pages 49–64. Academic
Press.
Galef, Jr., B. G. (2013). Imitation in animals: History, def-
inition, and interpretation of data from the psycholog-
ical laboratory. In Social learning, pages 15–40. Psy-
chology Press.
Gerull, F. C. and Rapee, R. M. (2002). Mother knows best:
effects of maternal modelling on the acquisition of
fear and avoidance behaviour in toddlers. Behaviour
Research and Therapy, 40(3):279–287.
Giraldeau, L.-A. (1997). The ecology of information use. In
Krebs, J. R. and Davies, N. B., editors, Behavioural
Ecology: An Evolutionary Approach, pages 42–68.
Blackwell Oxford, UK, 4 edition.
Hamid, O., Dautenhahn, K., and Nehaniv, C. L. (2020). En-
gineering social learning mechanisms for minimalistic
multi-agent robots. In 2020 3rd International Con-
ference on Control and Robots (ICCR), pages 90–99.
IEEE.
Heyes, C. M. (1994). Social learning in animals: categories
and mechanisms. Biological Reviews, 69(2):207–231.
Hoppitt, W. and Laland, K. N. (2013). Social learning:
an introduction to mechanisms, methods, and models.
Princeton University Press.
Lynch, T. and Martins, N. (2015). Nothing to fear? an anal-
ysis of college students’ fear experiences with video
games. Journal of Broadcasting & Electronic Media,
59(2):298–317.
MacInnis, C. C., Boss, H. C., and Bourdage, J. S. (2020).
More evidence of participant misrepresentation on
mturk and investigating who misrepresents. Person-
ality and Individual Differences, 152:109603.
Mineka, S. and Cook, M. (2013). Social learning and the
acquisition of snake fear in monkeys. In Social learn-
ing, pages 63–86. Psychology Press.
Nehaniv, C. L. and Dautenhahn, K. (2001). Like me?-
measures of correspondence and imitation. Cybernet-
ics & Systems, 32(1-2):11–51.
Palameta, B. and Lefebvre, L. (1985). The social transmis-
sion of a food-finding technique in pigeons: what is
learned? Animal Behaviour, 33(3):892–896.
Previde, E. P. and Poli, M. D. (1996). Social learning in
the golden hamster (mesocricetus auratus). Journal
of Comparative Psychology, 110(2):203.
Reeves, B. and Nass, C. (1996). The media equation: How
people treat computers, television, and new media like
real people. Cambridge University Press.
Seyfarth, R. M., Cheney, D. L., and Marler, P. (1980). Mon-
key responses to three different alarm calls: evidence
of predator classification and semantic communica-
tion. Science, 210(4471):801–803.
Steels, L. and Kaplan, F. (2000). Aibo’s first words: The
social learning of language and meaning. Evolution of
Communication, 4(1):3–32.
Terkel, J. (1996). Cultural transmission of feeding behavior
in the black rat (rattus rattus). In Heyes, C. M. and
B. G. Galef, J., editors, Social Learning in Animals:
The Roots of Culture, pages 17–47. Academic Press.
Tomasello, M. (2009). The Cultural Origins of Human Cog-
nition. Harvard University Press.
Zentall, T. R. (2001). Imitation in animals: evidence, func-
tion, and mechanisms. Cybernetics & Systems, 32(1-
2):53–96.
ICAART 2022 - 14th International Conference on Agents and Artificial Intelligence
372
