
NCBI Database on Cycloartenol Synthase
Mohammad Basyuni
1,2
, Rahmah Hayati
1
, Yuntha Bimantara
1
, Rizka Amelia
1
, Sumaiyah
3
, Era
Yusraini
4
, and Hirosuke Oku
5
1
Department of Forestry, Faculty of Forestry, Universitas Sumatera Utara, Jl. Tri Dharma Ujung No. 1 Medan, North
Sumatera 20155, Indonesia
2
Mangrove and Bio-Resources Group, Center of Excellence for Natural Resources Based Technology, Universitas
Sumatera Utara, Medan North Sumatera 20155, Indonesia.
3
Faculty of Pharmacy, Universitas Sumatera Utara, Medan 20155, Indonesia
4
Faculty of Agriculture, Universitas Sumatera Utara, Medan 20155, Indonesia
3
Molecular Biotechnology Group, Tropical Biosphere Research Center, University of the Ryukyus, 1 Senbaru, Nishihara,
Okinawa 903-0213, Japan
Keywords: Abiotic stress, cycloartenol, isoprenoid, oxidosqualene
Abstract: Cycloartenol synthase (EC 5.4.99.8) is a cycloartenol-converting enzyme. The current research describes
the search of cycloartenol synthase databases from the National Center for Biotechnology Information
(NCBI). A amount of precious information was generated by NCBI database search
(https:/www.ncbi.nlm.nih.gov/). Results discovered in 22 cycloartenol synthase databases. All literature,
genes, genetics, protein, genomes, and chemical features of cycloartenol synthase databases. Bookshelf,
MeSH (Medical Subject Headings) and PubMed Central were discussed in the literature. Gene was made up
of profiles from Gene, Gene Expression Omnibus (GEO), HomoloGene, PopSet, and UniGene. Data on
genetics such as MedGen was available for cycloartenol synthase. Proteins characteristic comprised
Conserved Domains, Identical groups of proteins, clusters of proteins, sparcle, and structure. Nucleotide and
sample were engaged in genomes. BioSystems, PubChem BioAssay, PubChem Compound, and PubChem
Substance are the chemical properties. The present work affords vital information in term biotechnology of
cycloartenol synthase.
1 INTRODUCTION
Phytosterol including cycloartenol is ordinary
chemical constituents in higher plants (Augustin et
al., 2011; Uddin et al., 2015). Phytosterols serve as
chemotaxonomy markers also have been shown
(Koch et al., 2011). Many phylogenetic analyses
have been suggested that plant oxidosqualene
cyclases (OSCs) are generally evolutionarily
derived from cycloartenol synthase (Zhang et al.,
2003; Basyuni et al., 2007a,b). More than
thousands different OSC genes together with
cycloartenol have been reported from plant
kingdom, which are expanded from a shared 2,3-
oxidosqualene substrate via OSCs, they separate in
the cyclization of this intermediacy into each of
phytosterol of cycloartenol and lanosterol or a
diversity of triterpenes (Augustin et al., 2011). In
plantae, cycloartenol synthase (CAS) along with
lanosterol synthase (LAS), family member of OSC
are accountable for biosynthesis of phytosterol,
and more OSCs are committed for triterpene
synthesis (Thimmappa et al., 2014).
Lanosterol and cycloartenol are significant
elements of the membrane and can provide
substrates for hormones of sterols (Bloch 1983).
Our previous studies have shown that membrane
lipids (triterpenoids and phytosterols) have an
evident role in mangrove modification to salinity
stress (Oku et al., 2003; Basyuni et al., 2009,
2012a,b), Saccharomyces cerevisiae, GIL77,
modulated the salt tolerance of lanosterol synthase
deficient (Inafuku et al., 2018). Even though a
significant number of studies in cycloartenol
synthases have been well documented (Agustin et
al., 2011; Xue et al., 2012; Gas-Pascual et al.,
2014; Moses et al., 2014; Thimmappa et al., 2014;),
Limited work concentrated on biotechnology data
from all accessible databases in cycloartenol
synthases. Here we report an alternative technique
16
Basyuni, M., Hayati, R., Bimantara, Y., Amelia, R., Sumaiyah, S., Yusraini, E. and Oku, H.
NCBI Database on Cycloartenol Synthase.
DOI: 10.5220/0008386800160019
In Proceedings of the International Conference on Natural Resources and Technology (ICONART 2019), pages 16-19
ISBN: 978-989-758-404-6
Copyright
c
2019 by SCITEPRESS – Science and Technology Publications, Lda. All rights reserved
to collect useful data needed in latest
biotechnology-related science studies on
cycloartenol synthase through a preferred search
engine. Therefore, the aim of this research was to
explain the implementation of the National Center
for Biotechnology Information (NCBI) databases
(GQuery) search to gain more deepful information
on updated cycloartenol synthesis biotechnology.
2 MATERIALS AND METHODE
The search engine for NCBI databases
(https:/www.ncbi.nlm.nih.gov/) was used to
produce a amount of useful rubber plant data
biotechnology. As stated previously on September
19, 2018, data bases were obtained by writing
Hevea brasiliensis in all database searches. All
databases composed of literature, health, genes,
protein, genomes, and chemical properties were
featured. Cycloartenol synthase (GQuery) was
performed. The bookshelf, MeSH (Medical
Subject Headings), NLM (National Medicine
Library) Catalog, PubMed, PubMed Central, EST,
Gene, GEO datasets, PopSet, Identical Protein
Groups, Protein, Sparde, Structure, Assembly,
BioProject, BioSample, Genome, GSS, Nucleotide,
Probe, SRA, Taxonomy, Biosystems, and
PubChem BioAssay were the data variables.
3 RESULTS AND DISCUSSIONS
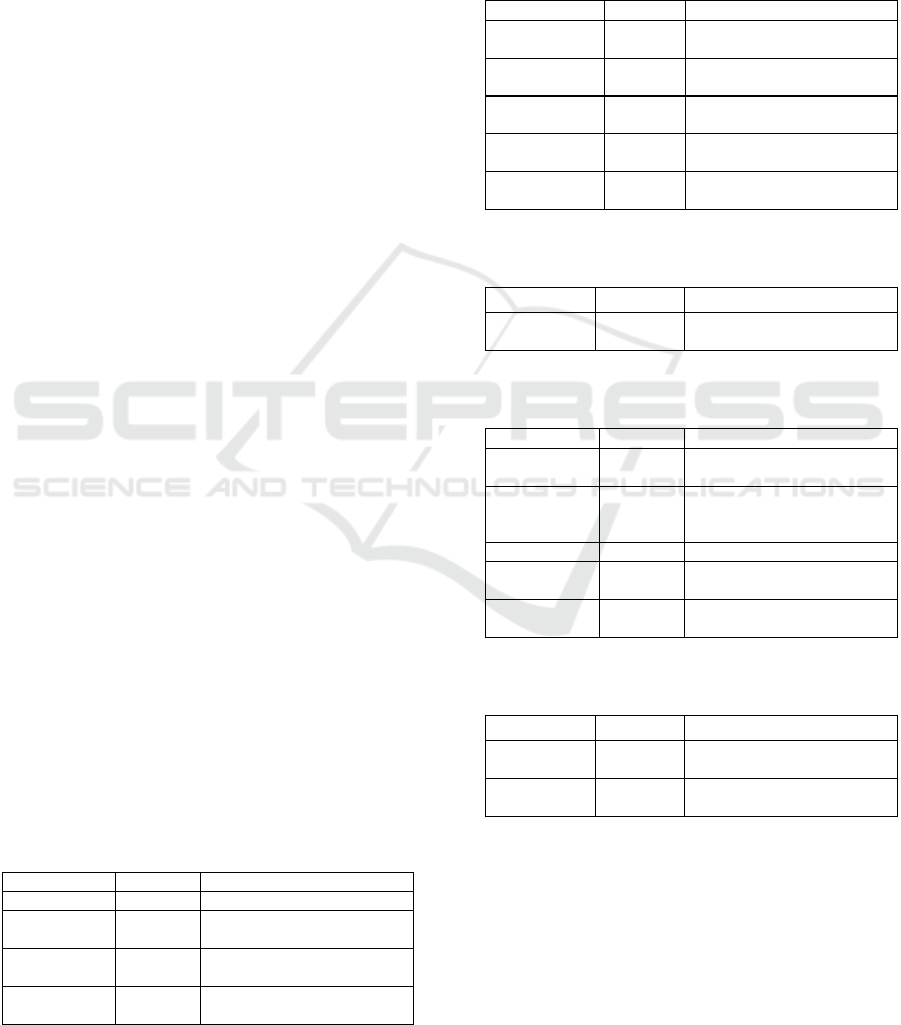
Table 1 demonstrates the literature on cycloartenol
synthase available in the NCBI. Four biographies
stored in countless numbers. The web NCBI
literature provides internet libraries and free access
to bookshelf data (two reports and books), 2 MeSH
(Medical Subject Headings), namely cycloartenol
synthase [Supplementary Concept] and 3-beta-(2-
(diethylamino) ethoxy) androst-5-en-17-one [
Additional Concept], ontology used to index
PubMed papers.
Table 1: Literature source NCBI database for
cycloartenol synthase
Literature
Total
Information
Bookshelf
2
Reports and books
Mesh
2
Medical topic in the
collections of the NLM
PubMed
312
Scientific and medical
abstracts/quotations
PubMed
Central
584
Articles in full-text journals
Furthermore, 87 Pubmed and 232 PubMed Central
documentations were stored as called the plant
cycloartenol synthase (Table 1). The PubMed
database includes quotes derived different topics in
many connections to reports of full-text papers on
rubber plants open access.
Table 2: Genes source NCBI database for cycloartenol
synthase
Health
Amount
Explanation
Gene
242
Gathered data about the gene
loci
GEO Profiles
116
Gene profile and molecular
abundance patterns
HomoloGene
2
Homologous gene sets for
chosen organisms
PopSet
3
Sets of phylogenetic and
population studies sequences
UniGene
66
Groups of transcripts
conveyed
Table 3: Genetics source NCBI database for cycloartenol
synthase
Genetics
Total
Information
MedGen
1
Literature and connections in
medical genetics
Table 4: Proteins source NCBI database for cycloartenol
synthase
Proteins
Number
Explanation
Conserved
Domains
1
Preserved protein domains
Identical
Protein
Groups
723
Sequences of proteins
classified by similarity
Protein
976
Sequences of proteins
Protein
Clusters
17
Sequence of protein clusters
based on similarity
Structure
2
Experimentally determined
biomolecular structures
Table 5: Genomes source NCBI database for Plant and
cycloartenol synthase
Genomes
Number
Description
Nucleotide
1583
RNA and DNA sequences
Probe
92
Biomolecular structures
experimentally determined
Information on genes sources is displayed in
Table 2. This data is as well as genes associated to
cycloartenol synthase than consisting of 242 genes
with top organisms are Rosa chinensis (15), Malus
domestica (14), Camellia sinensis (11), Zea mays
(10), Pyrus x bretschneideri (10), Gossypium
hirsutum (10), G. raimondii (10), Vitis vinifera
(10), Aegilops tauschii (8), Arachis hypogaea (8),
NCBI Database on Cycloartenol Synthase
17

Brachypodium distachyon (7), Phoenix dactylifera
(7), Nicotiana tabacum (7), and all other taxa.
Table 2 shows there is 116 GEO profiles, one
homologene, three popset, and 66 uniges. It is
important to note that top organisms for unigene
are Malus domestica (5), Zea mays (5), Panicum
virgatum (4), Brassica napus (3), Arabidopsis
thaliana (3), Citrus sinensis (3), Medicago
truncatula (3), Vitis vinifera (3), Triticum aestivum
(3), and Pinus taeda (3). The three popsets are
Nicotiana tabacum cycloartenol synthase (CAS1)
mRNA, complete cds, Bacillariophyta cycloartenol
synthase gene, partial cds, and Avena cycloartenol
synthase gene, complete cds.
Table 3 shows one genetics information on
medical genetics literature and links. On the other
hand, Table 4 depicts protein source from NCBI
database for there are five sources for protein that
consists of one conserved domain, 723 identical
protein groups, 976 proteins, 17 protein clusters,
and two structures. The conserved domains
comprise C-terminal domain of squalene-hopene
cyclase. EC:5.4.99.17 registered as squalene-
hopene cyclase, in bacteria activates the cyclisation
of squalene into hopene. This response is partly
cationic cyclisation cascade, to be similar to a
critical level in cholesterol biosynthesis. The C-
terminal half of the molecule belongs to this family
(Marchler-Bauer et al., 2016).
The same protein groups combined from 13
invertebrates, 437 plants and fungi, and 26
prokaryotes. It is interesting to note that 976
proteins are consisting of animals (2), plants (797),
fungi (1), protists (85), bacteria (90) and archaea
(1). Top organism for plants are 51 Oryza sativa
(International Rice Genome Sequencing Project,
2005), 46 Vitis vinifera (Roach et al., 2018) and
46 Zea mays (Soderlund et al., 2009), 21 Medicago
truncatula (Young et al., 2011), and 20
Arabidopsis thaliana (Corey et al., 1993).
The two structures of cycloartenol are structure
of human OSC in complex with Ro 48-8071
[Isomerase, EC: 5.4.99.7] and Human OSC
structure in lanosterol complex [Isomerase, EC:
5.4.99.7].
Table 5 displays 1583 nucleotides that
comprising of Animals(6), Plants(1,294), Fungi(1),
Protists(111), Bacteria(171), Archaea(1). The top
plant species as following Oryza sativa (55),
Malus domestica (45), Zea mays (43), Glycine
max (27), Vitis vinifera (27), and all other taxa
(1097). There are 92 probes in the cycloartenol
synthase. Table 6 describes the chemical source of
the NCBI database that consists of 10234
Biosystems, 107 PubChem bioassay, 22 PubChem
compound, and 58 PubChem substance.
Table 6: Chemicals source NCBI database for
cycloartenol synthase
Chemicals
Total
Information
BioSystems
10234
Molecular pathways
associated to genes, proteins
and chemicals
PubChem
BioAssay
107
Bioactivity screening
PubChem
Compound
22
Chemical data with systems,
experience and relationships
PubChem
Substance
58
Substance stored and
chemical data
Protein modeling of cycloartenol synthase from
mangrove trees has recently been defined (Basyuni
et al. 2018). The protein modeling for KcCAS in
Kandelia candel and RsCAS of Rhizophora stylosa
cycloartenol synthases was analyzed by Pyre2
showed homolous results with lightly divergent in
sequence similarity. By disparity, the Swiss model
for KcCAS had marginally greater sequence
homology (47.3%) and differentiated Qmean
(0.70) from RsCAS.
4 CONCLUSIONS
The web NCBI defines multiple biological and
biotechnology data on cycloartenol synthase. The
current work motivated scientists in the field of
biotechnology to obtain more advantages using the
NCBI search engine. The present study also
delivers crucial data regarding biotechnology of
triterpene synthases.
ACKNOWLEDGMENTS
This work was in part assisted by an International
Research Collaboration and Scientific Publication
Grant 2019 from Universitas Sumatera Utara.
REFERENCES
Augustin, J.M., Kuzina, V., Andersen, S.B. and Bak, S.,
2011. Molecular activities, biosynthesis and
evolution of triterpenoid saponins. Phytochemistry,
72, 435-457.
Basyuni, M., Oku, H., Tsujimoto, E., Kinjo, K., Baba, S.
and Takara, K., 2007a. Triterpene synthases from the
ICONART 2019 - International Conference on Natural Resources and Technology
18

Okinawan mangrove tribe, Rhizophoraceae. The
FEBS Journal, 274(19), 5028-5042.
Basyuni, M., Oku, H., Tsujimoto, E., and Baba, S. 2007b.
Cloning and functional expression of cycloartenol
synthases from mangrove species Rhizophora
stylosa Griff. and Kandelia candel (L.) Druce.
Bioscience, Biotechnology, and Biochemistry, 71(7),
1788-1792.
Basyuni, M., Baba, S., Inafuku, M., Iwasaki, H., Kinjo,
K. and Oku, H., 2009. Expression of terpenoid
synthase mRNA and terpenoid content in salt
stressed mangrove. Journal of Plant Physiology,
166(16), 1786-1800.
Basyuni, M., Baba, S., Kinjo, Y., Putri, L.A., Hakim, L.
and Oku, H., 2012a. Salt-dependent increase in
triterpenoids is reversible upon transfer to fresh
water in mangrove plants Kandelia candel and
Bruguiera gymnorrhiza. Journal of Plant
Physiology, 169(18), 1903-1908.
Basyuni, M., Baba, S., Kinjo, Y., and Oku, H. 2012b.
Salinity increases the triterpenoid content of a salt
secretor and a non-salt secretor mangrove. Aquatic
Botany, 97(1), 17-23.
Basyuni, M., Sulistiyono, N., Wati, R., Oku, H., Baba, S.,
and Sagami, H. 2018. Predicted cycloartenol
synthase protein from Kandelia obovata and
Rhizophora stylosa using online software of Phyre2
and Swiss-model. Journal of Physics : Conference
Series, 978 (1), 012077.
Bloch, K. E. 1983. Sterol, structure and membrane
function. Critical Reviews in Biochemistry, 14(1),
47-92.
Corey, E. J., Matsuda, S. P., and Bartel, B. 1993.
Isolation of an Arabidopsis thaliana gene encoding
cycloartenol synthase by functional expression in a
yeast mutant lacking lanosterol synthase by the use
of a chromatographic screen. In Proceedings of the
National Academy of Sciences, 90(24), 11628-11632.
Gas-Pascual, E., Berna, A., Bach, T. J., and Schaller, H.
2014. Plant oxidosqualene metabolism: cycloartenol
synthase–dependent sterol biosynthesis in Nicotiana
benthamiana. PLoS One, 9(10), e109156.
Inafuku, M., Basyuni, M., and Oku, H. 2018.
Triterpenoid modulates the salt tolerance of
lanosterol synthase deficient Saccharomyces
cerevisiae, GIL77. Saudi Journal of Biological
Sciences, 25(1), 1-9.
International Rice Genome Sequencing Project 2005.
The map-based sequence of the rice genome. Nature,
436(7052), 793.
Koch, B. P., Souza Filho, P. W., Behling, H., et al., 2011.
Triterpenols in mangrove sediments as a proxy for
organic matter derived from the red mangrove
(Rhizophora mangle). Organic Geochemistry, 42(1),
62-73.
Marchler-Bauer, A., Bo, Y., Han, L., et al., 2016.
CDD/SPARCLE: functional classification of
proteins via subfamily domain architectures. Nucleic
acids research, 45(D1), D200-D203.
Moses, T., Papadopoulou, K.K. and Osbourn, A., 2014.
Metabolic and functional diversity of saponins,
biosynthetic intermediates and semi-synthetic
derivatives. Critical reviews in biochemistry and
molecular biology, 49(6), pp.439-462.
Roach, M. J., Johnson, D. L., Bohlmann, J., et al., 2018.
Population sequencing reveals clonal diversity and
ancestral inbreeding in the grapevine cultivar
Chardonnay. PLoS Genetics, 14(11), e1007807.
Soderlund, C., Descour, A., Kudrna, D., et al., 2009.
Sequencing, mapping, and analysis of 27,455 maize
full-length cDNAs. PLoS Genetics, 5(11), e1000740.
Thimmappa, R., Geisler, K., Louveau, T., O'Maille, P.
and Osbourn, A., 2014. Triterpene biosynthesis in
plants. Annual Review of Plant Biology, 65, pp.225-
257.
Uddin, M. S., Sarker, M. Z. I., Ferdosh, S., Akanda, M. J.
H., Easmin, M. S., Bt Shamsudin, S. H., & Yunus, K.
B. (2015). Phytosterols and their extraction from
various plant matrices using supercritical carbon
dioxide: a review. Journal of the Science of Food
and Agriculture, 95(7), 1385-1394.
Xue, Z., Duan, L., Liu, D., et al., 2012. Divergent
evolution of oxidosqualene cyclases in plants. New
Phytologist, 193(4), 1022-1038.
Young, N. D., Debellé, F., Oldroyd, G. E., et al., 2011.
The Medicago genome provides insight into the
evolution of rhizobial symbioses. Nature, 480(7378),
520.
NCBI Database on Cycloartenol Synthase
19
