
Temporal Complexity of a Hopfield-Type Neural Model in Random and
Scale-Free Graphs
Marco Cafiso
1,2 a
and Paolo Paradisi
2,3 b
1
Department of Physics ’E. Fermi’, University of Pisa, Largo Bruno Pontecorvo 3, I-56127, Pisa, Italy
2
Institute of Information Science and Technologies ‘A. Faedo’, ISTI-CNR, Via G. Moruzzi 1, I-56124, Pisa, Italy
3
BCAM-Basque Center for Applied Mathematics, Alameda de Mazarredo 14, E-48009, Bilbao, Basque Country, Spain
fi
Keywords:
Bio-Inspired Neural Networks, Temporal Dynamics, Self-Organization, Connectivity, Intermittency,
Complexity.
Abstract:
The Hopfield network model and its generalizations were introduced as a model of associative, or content-
addressable, memory. They were widely investigated both as an unsupervised learning method in artificial
intelligence and as a model of biological neural dynamics in computational neuroscience. The complex-
ity features of biological neural networks have attracted the scientific community’s interest for the last two
decades. More recently, concepts and tools borrowed from complex network theory were applied to artificial
neural networks and learning, thus focusing on the topological aspects. However, the temporal structure is
also a crucial property displayed by biological neural networks and investigated in the framework of systems
displaying complex intermittency. The Intermittency-Driven Complexity (IDC) approach indeed focuses on
the metastability of self-organized states, whose signature is a power-decay in the inter-event time distribu-
tion or a scaling behaviour in the related event-driven diffusion processes. The investigation of IDC in neural
dynamics and its relationship with network topology is still in its early stages. In this work, we present the
preliminary results of an IDC analysis carried out on a bio-inspired Hopfield-type neural network comparing
two different connectivities, i.e., scale-free vs. random network topology. We found that random networks
can trigger complexity features similar to that of scale-free networks, even if with some differences and for
different parameter values, in particular for different noise levels.
1 INTRODUCTION
The Hopfield model is the first example of a recur-
rent neural network defined by a set of linked two-
state McCulloch-Pitts neurons evolving in discrete
time. The Hopfield model is similar to the Ising model
(Ernst, 1925) describing the dynamics of a spin sys-
tem in a magnetic field, but with all-to-all connec-
tivity among neurons instead of local spin-spin in-
teractions. More importantly, in his milestone pa-
per (Hopfield, 1982), Hopfield first introduced a rule
for changing the topology of network connections
based on external stimuli. Hopfield first proposed
and investigated the properties of this network model
in (Hopfield, 1982) and in successive works (Hop-
field, 1984; Hopfield, 1995). In particular, he also
proposed an extension of the original 1982 model
a
https://orcid.org/0009-0004-9519-1221
b
https://orcid.org/0000-0002-1036-4583
to a continuous-time leaky-integrate-and-fire neuron
model (Hopfield, 1984), also considering the case of
neurons with graded response, i.e., with a sigmoid
function mediating the voltage inputs from upstream
neurons. The main property of the Hopfield model
is that the connectivity matrix is allowed to change
according to a Hebbian rule (Hebb, 1949), which is
often summarised in the statement: “(neuron) cells
that fire together wire together” (L
¨
owel and Singer,
1992). To our knowledge, with this rule, the Hopfield
neural network model results in being the first model
used for the investigation of associative, or content-
addressable, memory. In the Artificial Intelligence
(AI) jargon, external stimuli correspond to the cases
of a training dataset that trigger the changes in the
connectivity matrix. This process involves decoding
the input data into a map of neural states (see, e.g.,
(Hopfield, 1995)).
The Hopfield neural model, as such and its varia-
tions, belong to the class of Spiking Neural Networks
438
Cafiso, M. and Paradisi, P.
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs.
DOI: 10.5220/0013007600003837
Paper published under CC license (CC BY-NC-ND 4.0)
In Proceedings of the 16th International Joint Conference on Computational Intelligence (IJCCI 2024), pages 438-448
ISBN: 978-989-758-721-4; ISSN: 2184-3236
Proceedings Copyright © 2024 by SCITEPRESS – Science and Technology Publications, Lda.

(SNNs). SSNs are still attracting considerable atten-
tion due to energy efficiency and high sensitivity to
temporal features of data and, even if they are nowa-
days less efficient concerning the classical deep neu-
ral networks, are thought to have great potential in
the context of neuromorphic computing (Davies et al.,
2018).
An interesting aspect studied by Hopfield is the
emergence of collective, i.e., self-organizing be-
haviour in relation to the stability of memories in the
network model. In this framework, Grinstein et al.
(Grinstein and Linsker, 2005) investigated the role
of topology in a neural network model extending the
Hopfield model to a more biologically plausible one,
but partially maintaining the computational advantage
of two-state McCulloch-Pitts neurons with respect to
continuous-time extension of the model. This was
achieved by introducing a maximum firing time and
a refractory time in the single neuron dynamics.
Since the last two decades, the interest towards the
complex topological features of neural networks has
gained momentum in many scientific fields involv-
ing concepts and tools of computational neuroscience
and/or AI (see, e.g., (Kaviani and Sohn, 2021) for a
survey). In particular, neural networks with complex
topologies, such as random (Erd ¨os-R
´
enyi) (Erd ¨os and
R
´
enyi, 1959), (Gros, 2013), small-world or scale-
free networks (Boccaletti et al., 2006), were shown
to outperform artificial neural networks with all-to-all
connectivity (McGraw and Menzinger, 2003; Torres
et al., 2004; Lu et al., 2006; Shafiee et al., 2016; Ka-
viani and Sohn, 2020; Adjodah et al., 2020; Kaviani
and Sohn, 2021).
Complexity is a general concept related to the
ability of a multi-component system to trigger self-
organizing behaviour, a property that is manifested
in the generation of spatio-temporal coherent states
(Paradisi et al., 2015; Grigolini, 2015; Paradisi and
Allegrini, 2017). Interestingly, in many research
fields, many authors consider the complexity of a sys-
tem as a concept essentially referring to its topolog-
ical structure, which is an approach borrowed from
graph theory and complex networks (Watts and Stro-
gatz, 1998; Barab
´
asi and Albert, 1999; Albert and
Barab
´
asi, 2002; Barab
´
asi and Oltvai, 2004). How-
ever, another aspect, which is often overlooked and
instead is typically a crucial feature of complex self-
organizing behaviour, is the temporal structure of the
system. This is intimately related not only to the
topological/geometrical structure of the network but
also to its dynamical properties, both at the level of
single nodes, of clusters of nodes, and as a whole.
Hereafter we refer to Temporal Complexity (TC),
or Intermittency-Driven Complexity (IDC), as the
property of the system to generate metastable self-
organized states, the duration of which is marked by
rapid transition events between two states (Grigolini,
2015). The underlying theories of TC/IDC refer
to Cox’s renewal theory (Cox, 1970), Cox’s failure
events are reinterpreted in a temporal sense, that is,
precisely as rapid transitions or jumps in the sys-
tem’s observed variables. The rapid transitions can
occur between two self-organized states or between a
self-organized state and a disordered or non-coherent
state. The sequence of transition events is then de-
scribed as a point process and the ideal condition for
TC/IDC is the renewal one (Cox, 1970; Bianco et al.,
2007; Paradisi et al., 2009), which is not easily de-
termined being mixed to spurious effects such as sec-
ondary events and noise (Paradisi and Allegrini, 2017;
Paradisi and Allegrini, 2015). The self-organizing be-
haviour can be detected by the recognition of given
patterns in the system’s variables, e.g., eddies in a tur-
bulent flow or synchronization epochs in neural dy-
namics, and the identification of events is achieved
by means of proper event detection algorithm in sig-
nal processing (Paradisi and Allegrini, 2017; Paradisi
and Cesari, 2023).
A general concept commonly accepted in the
complex system research field is that complexity fea-
tures are related to the power-law behaviour of some
observed features, e.g.: space and/or time correlation
functions, the distribution of some variables such as
the sequence of inter-event times and the size of neu-
ral avalanches (Beggs and Plenz, 2003). In TC/IDC a
crucial feature to be evaluated is the probability den-
sity function (PDF) of inter-event times, or Waiting
Times (WT). However, the WT-PDF is often blurred
by secondary events related to noise or other side ef-
fects (Allegrini et al., 2010; Paradisi and Allegrini,
2015). A more reliable analysis relies on diffusion
processes derived by the sequence of events and on
their scaling analysis (Akin et al., 2009).
This approach involves several scaling analyses
widely investigated in the literature that were inte-
grated in the Event-Driven Diffusion Scaling (ED-
DiS) algorithm (Paradisi and Allegrini, 2017; Paradisi
and Allegrini, 2015). This approach was also success-
fully applied in the context of brain data, being able to
characterize different brain states from wake, relaxed
condition to the different sleep stages (Paradisi et al.,
2013; Allegrini et al., 2013; Allegrini et al., 2015).
At present, the relationships between network
connectivity and temporal complexity by one hand
and the complexity of connectivity matrix and learn-
ing efficiency are still not clear. In this work, we
present some preliminary results regarding the first
aspect, having in mind the potential applications
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs
439

regarding the second aspect, i.e., connectivity vs.
learning efficiency. Along this line, an IDC anal-
ysis is carried out on a bio-inspired Hopfield-type
neural network comparing two different connectivi-
ties, i.e., scale-free vs. random network topology.
In Section 2 we introduce the methods to generate
the network topologies and the details of the bio-
inspired Hopfield-type neural network model. Sec-
tion 3 briefly describes the event-based scaling anal-
yses. In Section 4 we describe the results of numeri-
cal simulations and of their IDC analyses that are dis-
cussed in Section 5. Finally, we sketch some conclu-
sions in Section 6.
2 MODEL DESCRIPTION
2.1 Network Topology: Scale-Free vs.
Erd ¨os-R
´
enyi
We here consider two types of networks, both with
N number of nodes
1
. Our networks are constrained
to have the same minimum number k
0
of links for
each neuron and the same average number of links
⟨k⟩. In both cases, self-loops and multiple directed
edges from one node to another are excluded, fol-
lowing the methodology outlined in (Grinstein and
Linsker, 2005). The first class of networks is that
of Scale-Free (SF) graphs, characterized by a power-
law node out-degree distribution. The probability of a
node i to have k
i
outgoing links is given by:
∀i = 1, ...,N : P
SF
(k
i
) =
m
k
α
i
(1)
m =
α − 1
k
(1−α)
0
− (N − 1)
(1−α)
(2)
For the construction of SF networks, we used a
power-law exponent α = 2.5.
The second class of networks is that of Erd ¨os-
R
´
enyi (ER) graphs, which are random graphs where
each pair of distinct nodes is connected with a prob-
ability p. In an ER network with N nodes and with-
out self-loops, the average degree is simply given by:
⟨k⟩
ER
= p
ER
(N − 1). Then, from the equality of de-
gree averages: ⟨k⟩
ER
= ⟨k⟩
SF
we simply derive:
p
ER
=
⟨k⟩
SF
N − 1
(3)
1
Let us recall that the degree of a node in the network is
the number of links of the node itself. In a directed network,
each node has an out-degree, given by the number of outgo-
ing links, and an in-degree, given by the number of ingoing
links.
The theoretical mean out-degree of the SF network is
approximated by the following formula:
⟨k⟩ ≃ m
k
2−α
0
− (N − 1)
2−α
α − 2
=
=
α − 1
α − 2
k
2−α
0
− (N − 1)
2−α
k
1−α
0
− (N − 1)
1−α
that is obtained by considering k as a continuous ran-
dom variable. However, due to the large variability
of SF degree distribution, different statistical samples
drawn from P
SF
can have very different mean outde-
grees. Thus, we have chosen to numerically evalu-
ate the mean out-degree associated with the sample
drawn from P
SF
and to use this value instead of the
theoretical one to define P
SF
.
The algorithm used to generate the two network
topologies is as follows:
(SF)(a) For each node i, choose the out-degree k
i
as
the nearest integer of the real number defined
by:
k
i
= (((N − 1)
(1−α)
− k
(1−α)
0
)ξ
i
+ k
(1−α)
0
)
1
1−α
(4)
being ξ a random number uniformly dis-
tributed in [0,1]. This formula is obtained by
the cumulative function method. The drawn
k
i
are within the range [k
0
,N − 1].
(b) Given k
i
for each node i, the target nodes
are selected by drawing k
i
integer numbers
{ j
i
1
,..., j
i
k
i
} uniformly distributed in the set
{1,..., i − 1,i + 1,..., N}.
(c) Finally, the adjacency or connectivity matrix
is defined as:
A
SF
i j
=
1 if j ∈ T
i
= { j
i
1
,..., j
i
k
i
}
0 otherwise
(5)
With this choice, the in-degree distribution
results in a mono-modal distribution similar
to a Gaussian distribution.
(ER)(a) From the adjacency matrix A
SF
i j
the actual
mean out-degree ⟨k⟩
SF
is computed.
(b) For each couple of nodes (i, j) with j ̸= i a
random number ξ
i, j
is drawn from a uniform
distribution in [0,1].
(c) Finally, the adjacency matrix is defined as:
A
ER
i j
=
1 if ξ
i, j
< p
ER
0 otherwise
(6)
where p
ER
is given by Eq. (3).
NCTA 2024 - 16th International Conference on Neural Computation Theory and Applications
440

2.2 The Grinstein Hopfield-Type
Network Model
Grinstein et al. (Grinstein and Linsker, 2005) mod-
ify the Hopfield network by adding three elements:
(i) a random endogenous probability of firing p
endo
for each node; (ii) a maximum firing duration, thanks
to which the activity of a node shuts down after t
max
time steps; (iii) a refractory period such that a node,
once activated and subsequently deactivated, must re-
main inactive for at least t
re f
consecutive time-steps.
Each neuron i has two states: S
i
= 0 (”not firing”)
and S
i
= 1 (”firing at maximum rate”). The weight of
link from j to i is given by J
i j
(Nonconnected neurons
have J
i j
= 0). The network is initialized at time t = 0
by randomly setting each neuron state S
i
(0) equal to
1 with a probability p
init
that we chose equal to the
endogenous firing probability. At each time step the
weighted input to node i is defined:
I
i
(t) =
∑
j
J
i j
S
j
(t) (7)
as in the Hopfield network dynamics. The state of
the node i evolves with t according to the following
algorithm:
1. If S
i
(τ) for all τ = t,t − 1,· · · ,t − t
max
+ 1, then
S
i
(t + 1) = 0 (maximum firing duration rule).
2. If S
i
(t) = 1 and S
i
(t + 1) = 0 then S(τ) = 0 for
(t + 1) < τ ≤ (t +t
re f
) (refractory period rule).
3. If neither rule 1 nor rule 2 applies, then
(a) If I
i
(t) ≥ b
i
then S
i
(t + 1) = 1;
(b) If I
i
(t) < b
i
then S
i
(t + 1) = 1 with a probability
equal to p
endo
otherwise S
i
(t + 1) = 0.
where b
i
is the firing threshold of neuron i.
In our study, unlike Grinstein and colleagues, the
link’s weights and the firing thresholds are taken uni-
formly throughout the network: J
i j
= J and b
i
= b.
3 EVENT-DRIVEN DIFFUSION
SCALING ANALYSIS
The diffusion scaling analysis is a powerful method
for scaling detection and, when applied to a se-
quence of transition events, can give useful informa-
tion on the underlying dynamics that indeed gener-
ate the events. The complete IDC analysis involves
the Event-Driven Diffusion Scaling (EDDiS) algo-
rithm (Paradisi and Allegrini, 2015; Paradisi and Al-
legrini, 2017) with the computation of three different
random walks generated by applying three walking
rules to the sequence of observed transition events
and the computation of the second moment scaling
and the similarity of the diffusion PDF. The gen-
eral idea is based on the Continuous Time Random
Walk (CTRW) model (Montroll, 1964; Weiss and Ru-
bin, 1983), where a particle moves move only at the
event occurrence times. Here we limit to the so-called
Asymmetric Jump (AJ) walking rule (Grigolini et al.,
2001), which simply consists of making a unitary
jump ahead when an event occurs and, thus, corre-
sponds to the counting process generated by the event
sequence:
X(t) = #{n : t
n
< t}. (8)
The method used to extract events from the sim-
ulated data and the scaling analyses are described in
the following.
3.1 Neural Coincidence Events
The IDC features here investigated are applied to
coincidence events that are defined as the events at
which a minimum number N
c
of neuron fires at the
same time. Then, given the global set of single neu-
ron firing times, the coincidence event time is defined
as the occurrence time of more than N
c
nodes fir-
ing simultaneously, i.e., in a tolerance time interval
of duration ∆t
c
. Hereafter we always set ∆t
c
equal
to a sampling time, i.e., ∆t
c
= 1, which is equiva-
lent to look for simultaneous events. The total ac-
tivity distribution of the network corresponds to the
size distribution of coincidences with minimum num-
ber N
c
= 1: P(n
c
|N
c
= 1). The actual threshold N
c
here applied is defined by computing the 35th per-
centile of P(n
c
|N
c
= 1). Then, each n-th coincidence
event is described by its occurrence time t
c
(n) and its
size S
c
(n).
3.2 Detrended Fluctuation Analysis
(DFA)
DFA is a well-known algorithm (see, e.g., (Peng et al.,
1994)) that is widely used in the literature for the eval-
uation of the second-moment scaling H defined by:
F
2
(∆t) = ⟨(∆X(∆t) − ∆X
trend
(∆t))
2
⟩ ∼ t
2H
(9)
F(∆t) = a · ∆t
H
⇒
⇒ log(F(∆t)) = log(a) + H · log(∆t) (10)
being ∆X(t,∆t) = X(t + ∆t) − X (t). We use the no-
tation H as this scaling exponent is essentially the
same as the classical Hurst similarity exponent (Hurst,
1951). X
trend
(∆t) is a proper local trend of the time
series. The DFA is computed over different values of
the time lag ∆t and the statistical average is carried
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs
441

out over a set of time windows of duration ∆t into
which the time series is divided. In the EDDiS ap-
proach, DFA is applied to different event-driven dif-
fusion processes (see, e.g., (Paradisi et al., 2012; Par-
adisi and Allegrini, 2017)). To check the validity of
Eq. (10) on the data and evaluate the exponent H is
it sufficient to carry out a linear best fit in the loga-
rithmic scale. To perform the DFA we employed the
function MFDFA of package MFDFA of Python (Ry-
din Gorj
˜
ao et al., 2022).
3.3 Diffusion Entropy
The Diffusion Entropy (DE) is defined as the Shannon
entropy of the diffusion process X(t) and was exten-
sively used in the scaling detection of complex time
series (Grigolini et al., 2001; Akin et al., 2009). The
DE algorithm is the following:
1. Given a time lag ∆t, split the time series X(t)
into overlapping time windows of duration ∆t and
compute: ∆X(t,∆t) = X (t + ∆t) − X (t), ∀ t ∈
[0,t − ∆t].
2. For each time lag ∆t, evaluate the distribution
p(∆x,∆t).
3. Compute the Shannon entropy:
S(∆t) = −
Z
+∞
−∞
p(∆x,∆t)log p(∆x,∆t)dx (11)
If the probability density function (PDF) is self-
similar, i.e., p(∆x,∆t) = f (∆x/∆t
δ
)/∆t
δ
, it re-
sults:
S(∆t) = A + δlog(∆t + T ) (12)
To check the validity of Eq. (12) on the data and
evaluate the exponent δ is it sufficient to carry out
a linear best fit with a logarithmic scale on the
time axis.
4 NUMERICAL SIMULATIONS
AND RESULTS
We performed a comprehensive parametric analysis
with a fixed t
max
value of 3, while systematically vary-
ing the other parameters. These include J which was
constrained to integer values ranging from 1 to 4,
p
endo
set to 0.001, 0.01 and 0.1, k
0
limited to integers
spanning from 1 to 5, b with two options of either 2
or 3, and t
re f
set at 0, 4, 6, 8 and 10. For dimen-
sional reasons, the model’s dynamics depend only on
the adimensional parameter π =
J
b
, which is conve-
niently used in the plots summarising the parametric
analysis, i.e. Figs. 1 and 2. The simulations were car-
ried out for 20000 time steps within networks com-
prising N = 1000 neurons. We analyzed the network
behaviours by examining two key metrics: the total
activity distribution P(n
c
|N
c
= 1) and the average ac-
tivity over time. The parametric analysis results are
reported in Fig. 1 and Fig. 2. For the ER networks
we have identified the following qualitative behaviors
in the total activity distributions:
1. Asymmetric Bell Curve distribution
2. Symmetric Bell Curve distribution
3. Bell curve and transition to multi-modal distribu-
tion
4. Cycle distribution
5. Mono-modal at zero distribution
6. Multi-modal distribution
7. Peak with Power-Law distribution
8. Peak with Power-Law and transition to multi-
modal distribution
9. Power-Law distribution
10. Power-Law with Multiple Peaks distribution
For the SF networks we have identified the follow-
ing qualitative behaviors in the total activity distribu-
tions:
1. Asymmetric Bell Curve distribution
2. Symmetric Bell Curve distribution
3. Cycle distribution
4. Mono-modal at zero distribution
5. Multi-modal distribution
6. Peak with Power-Law distribution
7. Peak with Power-Law and transition to multi-
modal distribution
8. Power-Law distribution
9. Power-Law and transition to cycle distribution
Figures 4 and 5 report the results for ER and SF
networks, respectively.
We selected specific cases for further investigation us-
ing TC/IDC analysis. Specifically, we analyzed the
WTs between successive coincidence events by ap-
plying the DFA and DE analyses. We report some
of the results in Fig. 3, where a few relevant cases
involving power-law behaviour were selected. The
best-fit values of the scaling exponents H and δ are
reported in Table 1.
NCTA 2024 - 16th International Conference on Neural Computation Theory and Applications
442
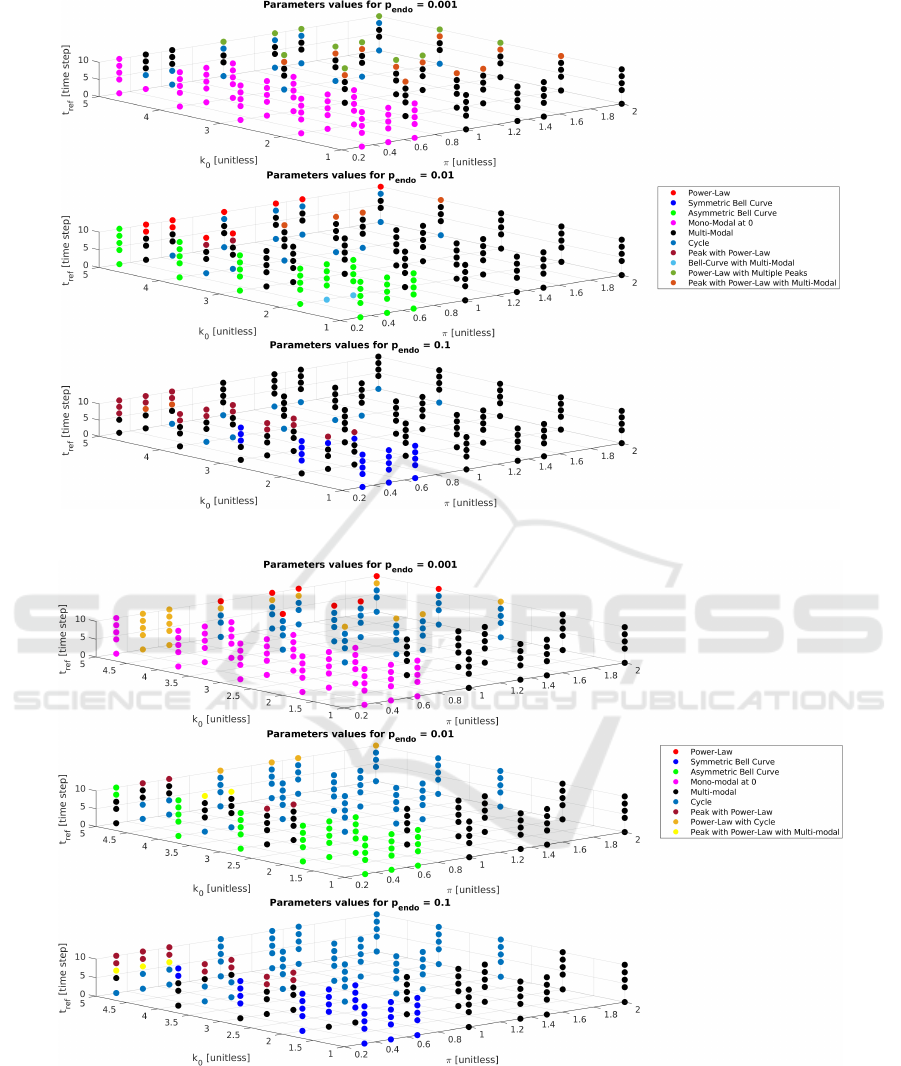
Figure 1: Results of parameter analysis derived from the behaviour of the total activity distribution in ER networks.
Figure 2: Results of parameter analysis derived from the behaviour of the total activity distribution in SF network.
5 DISCUSSION
Our numerical simulations showed a large variety of
behaviours in the Hopfield-type network in both net-
work topologies, i.e., random (ER) and scale-free
(SF). As can be seen from Figs. 4 and 5, most quali-
tative behaviors can be found in both network topolo-
gies, but with different sets of parameters. However,
some differences can be seen: (i) “bell curve with
multimodal” and “power-law with multiple peaks”
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs
443

Table 1: Result of fits for DFA and DE. For the “Power-Law with Cycle (SF)” row: Power-Law part is indicated with (PL), and
Cycle part is indicated with (C). In the “Power-Law (SF)” case, the fit δ = 0.184 for DE in the time range ∆t ∼ 10
3
− 3 · 10
3
was not reported.
DFA (H) DE (δ)
Short-Time Long-Time Short-Time Long-Time
Power-Law (ER) 0.061 1.164 / 0.352
Power-Law (SF) 0.070 1.061 / 0.382
Power-Law with Multiple Peaks (ER) 0.084 1.075 / 0.407
Power-Law with Cycle (SF) 0.105 (PL) 1.138 (PL) / /
0.078 (C) 1.327 (C) / /
(a) DFA (b) DE
Figure 3: DFA and DE analyses. Panels (1) and (2) same parameter set k
0
= 5, t
re f
= 10, b = 2, J = 3 and p
endo
= 0.01 (same
as panels (9) of Fig. 4 and (9) of Fig. 5). Panels (3) and (4) same as before but with different noise level p
endo
= 0.001 (same
as panels (10) of Fig. 4 and (8) of Fig. 5).
behaviours appear only in the ER network topology
(panels (3) and (10) in Fig. 4), while (ii) “power-
law with cycle” behaviour is exclusive of the SF net-
work topology (panels (9) of Fig. 5). Interestingly, all
other behaviours are seen in both topologies, even if
with slight differences. In particular, the emergence
of power-law behaviour in the total activity distribu-
tion (compare, e.g., panel (9) of Fig. 4 with panel
(8) of Fig. 5). It is worth noting that different kinds
of power-law behaviours are seen in both topologies
(panels (7-10) of Fig. 4 and panels (6-9) of Fig. 5).
As can be seen also from Figs. 1 and 2, for the pure
power-law behaviour (red dots) the main difference
seems to lie in the different noise levels: p
endo
= 0.01
for ER and p
endo
= 0.001 for SF.
Another remarkable observation regards the abrupt
transition among different behaviours seen in some
specific cases. In particular, some cases display a ini-
tial power-law behaviour that can persist for a very
long time, but then it is followed by an unexpected
transition to multimodal or cycle behaviour (panels
(8) in Fig. 4 and panels (7) and (9) in Fig. 5). Only
in ER networks, it is also seen a transition between
a mono-modal to a multi-modal distribution where
maxima are shifted towards higher values of total ac-
tivity (panels (3) of Fig. 4 ).
Fig. 3 shows some relevant behaviours in both DE
and DFA functions. In particular, we chose to an-
alyze: (i) pure power-law behavior in both topolo-
gies (panels (1) and (4)), which surprisingly arises
for similar parameter sets apart from the noise level;
(ii) power-law with multiple peaks in ER (panel (3))
and (iii) power-law with a transition to a cycle for SF
(panel (2)). Regarding “power-law with cycle” (panel
(2)), due to the rapid transition from the power-law
to the cycle behaviour, DFA and DE were applied
separately to the two regimes. Surprisingly, the cy-
cle regimes gives a pattern of DFA and DE similar to
that of the power-law regime, even if with different
slopes. Reliable fit values for H and δ are reported
in Table 1. It can be seen that the qualitative be-
haviour of DFA are essentially the same in the differ-
NCTA 2024 - 16th International Conference on Neural Computation Theory and Applications
444

(a)
(b)
Figure 4: (a) Average Activity plots over time and (b) Histograms of Total Activity for all the qualitative behaviours found in
ER networks. Parameters for each panel: (1) k
0
= 1, t
re f
= 4, b = 2, p
endo
= 0.01, and J = 1; (2) k
0
= 1, t
re f
= 4, b = 2, p
endo
=
0.1, and J = 1; (3) k
0
= 2, t
re f
= 0, b = 2, p
endo
= 0.01, and J = 1; (4) k
0
= 4, t
re f
= 0, b = 2, p
endo
= 0.01, and J = 1; (5) k
0
=
1, t
re f
= 4, b = 2, p
endo
= 0.001, and J = 1; (6) k
0
= 3, t
re f
= 6, b = 3, p
endo
= 0.1, and J = 2; (7) k
0
= 5, t
re f
= 6, b = 3, p
endo
=
0.1, and J = 1; (8) k
0
= 5, t
re f
= 6, b = 3, p
endo
= 0.1, and J = 2; (9) k
0
= 5, t
re f
= 10, b = 2, p
endo
= 0.01, and J = 3; (10) k
0
=
5, t
re f
= 10, b = 2, p
endo
= 0.001, and J = 3.
ent cases. All the investigated cases have the same pa-
rameters, except for the noise level, which is given by
p
endo
= 0.01 for the top panels and 0.001 for the bot-
tom panels. In summary, we have: (i) short-time with
very low H, associated with highly anti-persistent cor-
relations; (ii) long-time with very high H ∼ 1, except
in panel (3) where H > 1, associated with highly per-
sistent correlations and superdiffusion. Interestingly,
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs
445

(a)
(b)
Figure 5: (a) Average Activity plots over time and (b) Histograms of Total Activity for all the qualitative behaviours found in
SF networks. Parameters for each panel: (1) k
0
= 1, t
re f
= 0, b = 2, p
endo
= 0.01, and J = 1; (2) k
0
= 1, t
re f
= 0, b = 2, p
endo
=
0.1, and J = 1; (3) k
0
= 5, t
re f
= 0, b = 3, p
endo
= 0.01, and J = 2; (4) k
0
= 1, t
re f
= 0, b = 2, p
endo
= 0.001, and J = 1; (5) k
0
=
3, t
re f
= 0, b = 3, p
endo
= 0.1, and J = 1; (6) k
0
= 3, t
re f
= 10, b = 3, p
endo
= 0.1, and J = 2; (7) k
0
= 5, t
re f
= 10, b = 3, p
endo
=
0.1, and J = 2; (8) k
0
= 5, t
re f
= 10, b = 2, p
endo
= 0.001, and J = 3; (9) k
0
= 5, t
re f
= 10, b = 2, p
endo
= 0.01, and J = 3.
we get H ≃ 1 for p
endo
= 0.001 in both topologies,
while the pure power-law, which occurs for different
noise levels in the two topologies gives a larger value
of H for the ER network (H ≃ 1.16). The DE dis-
plays a power-law only in the long-time regime that
is, at variance with the DFA, in agreement with a sub-
diffusive behaviour. This is not directly related to the
persistence of correlations, but directly to the shape
of the diffusion PDF. In summary, in the long-time
regime of time lags, the diffusion generated by the
coincidence events, which are a manifestation of self-
organizing behaviour, shows highly persistent corre-
lations, as revealed by DFA, associated with a subd-
iffusive behaviour in the DE analysis. This could be
compatible with a very slow power-law decay in the
WT-PDF, i.e., ψ(τ) ∼ τ
µ
with µ < 2.
NCTA 2024 - 16th International Conference on Neural Computation Theory and Applications
446

6 CONCLUDING REMARKS
Here we have investigated a Hopfield-type model
and, in particular, the model proposed in (Grinstein
and Linsker, 2005), being a simple prototype of bio-
inspired neural model. This includes bio-inspired
features, such as the refractory time and the maxi-
mum firing time that can be tuned and interpreted,
in the context of AI, as hyper-parameters. A partic-
ularly interesting bio-inspired feature is also encoded
in the learning mechanism of Hopfield-type networks,
which is exactly the Hebbian bio-inspired, unsuper-
vised, learning. The network topology is recognized
to also play a central role in both global network dy-
namics and learning efficiency. This last feature is of
particular interest in the AI field.
In particular, some authors focused on the effect
of topological structure on learning features (Kaviani
and Sohn, 2021), in some cases finding a better learn-
ing performance associated with specific topologies,
e.g., scale-free and/or small-world (Lu et al., 2023).
The present work represents a preliminary investiga-
tion of the relationships among connectivity features
and temporal complexity of a simple spiking neural
network without learning algorithms. Interestingly,
different topological structures can give similar dy-
namical behaviours and complexity features (see, e.g,
Fig. 4b, panel (9), with Fig. 5b, panel (8), and Fig.
3a, panels (1) and (4)).
Regarding the relationship between connectivity
and temporal complexity, further investigations are
needed to better understand the reason why very dif-
ferent topologies can give similar complexity. We ex-
pect these further investigations to deepen the under-
standing of the relationship between dynamical fea-
tures of the network, e.g., temporal complexity, con-
nectivity structure and learning features, such as stor-
age capacity.
We also plan future investigations regarding the re-
lationship between connectivity structure and learn-
ing algorithms, to study how the performance mea-
sures, jointly with the evaluation of complexity in-
dices, change with the number of stored patterns (e.g.,
in the Hopfield model).
7 FUNDING
This work was supported by the Next-Generation-
EU programme under the funding schemes PNRR-
PE-AI scheme (M4C2, investment 1.3, line on AI)
FAIR “Future Artificial Intelligence Research”, grant
id PE00000013, Spoke-8: Pervasive AI.
REFERENCES
Adjodah, D., Calacci, D., Dubey, A., Goyal, A., Krafft,
P., Moro, E., and Pentland, A. (2020). Leveraging
communication topologies between learning agents in
deep reinforcement learning. volume 2020-May, page
1738 – 1740.
Akin, O., Paradisi, P., and Grigolini, P. (2009).
Perturbation-induced emergence of poisson-like be-
havior in non-poisson systems. J. Stat. Mech.: Theory
Exp., page P01013.
Albert, R. and Barab
´
asi, A.-L. (2002). Statistical mechanics
of complex networks. Reviews of Modern Physics,
74(1):47 – 97.
Allegrini, P., Menicucci, D., Bedini, R., Gemignani, A., and
Paradisi, P. (2010). Complex intermittency blurred
by noise: theory and application to neural dynamics.
Phys. Rev. E, 82(1 Pt 2):015103.
Allegrini, P., Paradisi, P., Menicucci, D., Laurino, M., Be-
dini, R., Piarulli, A., and Gemignani, A. (2013). Sleep
unconsciousness and breakdown of serial critical in-
termittency: New vistas on the global workspace.
Chaos Solitons Fract., 55:32–43.
Allegrini, P., Paradisi, P., Menicucci, D., Laurino, M., Pi-
arulli, A., and Gemignani, A. (2015). Self-organized
dynamical complexity in human wakefulness and
sleep: Different critical brain-activity feedback for
conscious and unconscious states. Phys. Rev. E Stat.
Nonlin. Soft Matter Phys, 92(3).
Barab
´
asi, A.-L. and Albert, R. (1999). Emergence of scal-
ing in random networks. Science, 286(5439):509 –
512.
Barab
´
asi, A.-L. and Oltvai, Z. N. (2004). Network biology:
Understanding the cell’s functional organization. Na-
ture Reviews Genetics, 5(2):101 – 113.
Beggs, J. M. and Plenz, D. (2003). Neuronal avalanches
in neocortical circuits. Journal of neuroscience,
23(35):11167–11177.
Bianco, S., Grigolini, P., and Paradisi, P. (2007). A fluctu-
ating environment as a source of periodic modulation.
Chem. Phys. Lett., 438(4-6):336–340.
Boccaletti, S., Latora, V., Moreno, Y., Chavez, M., and
Hwang, D.-U. (2006). Complex networks: Structure
and dynamics. Phys. Rep., 424(4-5):175–308. DOI:
10.1016/j.physrep.2005.10.009.
Cox, D. (1970). Renewal Processes. Methuen & Co., Lon-
don. ISBN: 0-412-20570-X; first edition 1962.
Davies, M., Srinivasa, N., Lin, T.-H., Chinya, G., Cao, Y.,
Choday, S. H., Dimou, G., Joshi, P., Imam, N., Jain,
S., Liao, Y., Lin, C.-K., Lines, A., Liu, R., Math-
aikutty, D., McCoy, S., Paul, A., Tse, J., Venkatara-
manan, G., Weng, Y.-H., Wild, A., Yang, Y., and
Wang, H. (2018). Loihi: A neuromorphic many-
core processor with on-chip learning. IEEE Micro,
38(1):82 – 99.
Erd ¨os, P. and R
´
enyi, A. (1959). On random graphs i. Pub-
licationes Mathematcae, 6(3-4):290–297.
Ernst, I. (1925). Beitrag zur theorie des ferromag-
netismus. Zeitschrift f
¨
ur Physik A Hadrons and Nu-
clei, 31(1):253–258.
Temporal Complexity of a Hopfield-Type Neural Model in Random and Scale-Free Graphs
447

Grigolini, P. (2015). Emergence of biological complexity:
Criticality, renewal and memory. Chaos Solit. Frac-
tals, 81(Part B):575–88.
Grigolini, P., Palatella, L., and Raffaelli, G. (2001). Asym-
metric anomalous diffusion: an efficient way to detect
memory in time series. Fractals, 9(04):439–449.
Grinstein, G. and Linsker, R. (2005). Synchronous neural
activity in scale-free network models versus random
network models. Biological Sciences, 102(28):9948–
9953.
Gros, C. (2013). Complex and adaptive dynamical systems:
A primer, third edition.
Hebb, D. O. (1949). The Organization of Behavior: A Neu-
ropsychological Theory. Wiley & Sons, New York.
Hopfield, J. (1982). Neural networks and physical systems
with emergent collective computational abilities. Pro-
ceedings of the National Academy of Sciences of the
United States of America, 79(8):2554 – 2558.
Hopfield, J. (1984). Neurons with graded response have
collective computational properties like those of two-
state neurons. Proceedings of the National Academy
of Sciences of the United States of America, 81(10
I):3088 – 3092.
Hopfield, J. (1995). Pattern recognition computation us-
ing action potential timing for stimulus representation.
Nature, 376(6535):33 – 36.
Hurst, H. (1951). Long-term storage capacity of reservoirs.
Trans. Am. Soc. Civil Eng., 116(1):770–799. DOI:
10.1061/TACEAT.0006518.
Kaviani, S. and Sohn, I. (2020). Influence of random topol-
ogy in artificial neural networks: A survey. ICT Ex-
press, 6(2):145 – 150.
Kaviani, S. and Sohn, I. (2021). Application of complex
systems topologies in artificial neural networks opti-
mization: An overview. Expert Systems with Applica-
tions, 180.
L
¨
owel, S. and Singer, W. (1992). Selection of intrinsic hor-
izontal connections in the visual cortex by correlated
neuronal activity. Science, 255(5041):209 – 212.
Lu, J., He, J., Cao, J., and Gao, Z. (2006). Topology influ-
ences performance in the associative memory neural
networks. Physics Letters A, 354(5-6):335 – 343.
Lu, L., Gao, Z., Wei, Z., and Yi, M. (2023). Working
memory depends on the excitatory–inhibitory balance
in neuron–astrocyte network. Chaos: An Interdisci-
plinary Journal of Nonlinear Science, 33(1):013127.
McGraw, P. N. and Menzinger, M. (2003). Topology and
computational performance of attractor neural net-
works. Physical Review E, 68(4 2):471021 – 471024.
Montroll, E. (1964). Random walks on lattices. Proc. Symp.
Appl. Math., 16:193–220.
Paradisi, P. and Allegrini, P. (2015). Scaling law of diffusiv-
ity generated by a noisy telegraph signal with fractal
intermittency. Chaos Soliton Fract, 81(Part B):451–
62.
Paradisi, P. and Allegrini, P. (2017). Intermittency-driven
complexity in signal processing. In Barbieri, R.,
Scilingo, E. P., and Valenza, G., editors, Complex-
ity and Nonlinearity in Cardiovascular Signals, pages
161–195. Springer, Cham.
Paradisi, P., Allegrini, P., Gemignani, A., Laurino, M.,
Menicucci, D., and Piarulli, A. (2013). Scaling and in-
termittency of brain events as a manifestation of con-
sciousness. AIP Conf. Proc, 1510:151–161. DOI:
10.1063/1.4776519.
Paradisi, P. and Cesari, R. (2023). Event-based complex-
ity in turbulence. Cambridge Scholar Publishing. In
Paolo Grigolini and 50 Years of Statistical Physics
(edited by B.J. West and S. Bianco), ISBN: 1-5275-
0222-8.
Paradisi, P., Cesari, R., Donateo, A., Contini, D., and Alle-
grini, P. (2012). Scaling laws of diffusion and time in-
termittency generated by coherent structures in atmo-
spheric turbulence. Nonlinear Proc. Geoph., 19:113–
126. P. Paradisi et al., Corrigendum, Nonlin. Processes
Geophys. 19, 685 (2012).
Paradisi, P., Cesari, R., and Grigolini, P. (2009). Superstatis-
tics and renewal critical events. Cent. Eur. J. Phys.,
7:421–431.
Paradisi, P., Kaniadakis, G., and Scarfone, A. M. (2015).
The emergence of self-organization in complex sys-
tems - preface. Chaos Soliton Fract, 81(Part B):407–
11.
Peng, C.-K., Buldyrev, S. V., Havlin, S., Simons, M., Stan-
ley, H. E., and Goldberger, A. L. (1994). Mosaic or-
ganization of dna nucleotides. Phys Rev E, 49:1685–
1689.
Rydin Gorj
˜
ao, L., Hassan, G., Kurths, J., and Witthaut, D.
(2022). Mfdfa: Efficient multifractal detrended fluc-
tuation analysis in python. Computer Physics Com-
munications, 273:108254.
Shafiee, M. J., Siva, P., and Wong, A. (2016). Stochasticnet:
Forming deep neural networks via stochastic connec-
tivity. IEEE Access, 4:1915 – 1924.
Torres, J., Mu
˜
noz, M., Marro, J., and Garrido, P. (2004).
Influence of topology on the performance of a neural
network. Neurocomputing, 58-60:229 – 234.
Watts, D. J. and Strogatz, S. H. (1998). Collective dynamics
of ’small-world9 networks. Nature, 393(6684):440 –
442.
Weiss, G. H. and Rubin, R. J. (1983). Random walks: the-
ory and selected applications. Advances in Chemical
Physics, 52:363–505.
NCTA 2024 - 16th International Conference on Neural Computation Theory and Applications
448
