
The Exhausted Brain Theory: An Energy-Based Framework for
Understanding Visually Induced Motion Sickness
Diego Monteiro
1 a
and Hai-Ning Liang
2 b
1
Immersion and Interaction Lab - LII, ESIEA, Laval, France
2
Computational Media and Arts Thrust, The Hong Kong University of Science and Technology (Guangzhou),
Guangzhou, China
Keywords:
Visually Induced Motion Sickness, Exhausted Brain Theory, Energy Metabolism, Sensory Conflict, Virtual
Reality, Predictive Coding, Computational Neuroscience.
Abstract:
Visually Induced Motion Sickness (VIMS) poses a persistent challenge in various scenarios, from virtual and
augmented reality (VR/AR) to transportation and simulation-based training. Existing theories, such as sensory
conflict and postural instability, offer partial insights but fail to fully explain the metabolic and cognitive
dynamics underlying VIMS. This paper introduces the Exhausted Brain Theory, which proposes that VIMS
arises from excessive energy demands on the brain as it recalibrates internal models to resolve conflicting
sensory inputs. Drawing from computational neuroscience, information theory, and energy metabolism, the
theory highlights how sensory conflicts overwhelm neural processing, deplete energy reserves, and disrupt
predictive coding mechanisms. We discuss implications for modeling, detection, and mitigation of VIMS,
including energy-efficient VR design, targeted acclimatization protocols, and personalized interventions. By
integrating diverse perspectives, this theory provides a unifying framework to advance understanding of VIMS
and guide future research on its prevention and management.
1 INTRODUCTION
In 1835, Charles Darwin said “. . . I continue to suf-
fer so much from sea-sickness, that nothing,[. . . ], can
make up for the misery. . . ” (Dobie, 2019) and he is
not alone. Motion sickness is also a problem that af-
fects millions of people (Dobie, 2019). In this posi-
tion paper, we use the term Visually Induced Motion
Sickness (VIMS) as an umbrella term to encompass
motion sickness, cybersickness, and simulator sick-
ness. VIMS has been a persistent challenge since the
advent of transportation and, more recently, virtual re-
ality (VR) technologies, including even 2D videos.
Despite centuries of study, a comprehensive under-
standing of its mechanisms remains elusive. Current
theories, such as the sensory conflict theory (Reason,
1978b) and the postural instability theory (Riccio and
Stoffregen, 1991), while valuable, fail to fully explain
all aspects of VIMS, particularly its variability across
individuals and situations.
VIMS manifests across a wide range of scenarios.
In transportation, it affects passengers in cars, boats,
a
https://orcid.org/0000-0002-1570-3652
b
https://orcid.org/0000-0003-3600-8955
airplanes, and even space vehicles, with symptoms
ranging from mild discomfort to severe nausea, and
the advent of autonomous vehicles introduces new
challenges as passengers become more disconnected
from vehicle control. In VR, augmented reality (AR),
and mixed reality (MR) applications, users often ex-
perience it during immersive experiences, limiting
the technology’s potential in fields such as education,
healthcare, and entertainment. VIMS affects mili-
tary personnel and civilian trainees using high-fidelity
simulators for aircraft, vehicles, and complex machin-
ery. Even in everyday scenarios, users of smartphones
and tablets can experience discomfort when viewing
motion-rich content or using navigation apps. As 3D
displays become more common in consumer electron-
ics, cinema, and gaming, a broader population is ex-
posed to potential visual discomfort and sickness.
Monteiro, D. and Liang, H.-N.
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness.
DOI: 10.5220/0013317000003912
Paper published under CC license (CC BY-NC-ND 4.0)
In Proceedings of the 20th International Joint Conference on Computer Vision, Imaging and Computer Graphics Theory and Applications (VISIGRAPP 2025) - Volume 1: GRAPP, HUCAPP
and IVAPP, pages 627-638
ISBN: 978-989-758-728-3; ISSN: 2184-4321
Proceedings Copyright © 2025 by SCITEPRESS – Science and Technology Publications, Lda.
627

2 THEORETICAL BACKGROUND
2.1 Visually Induced Motion Sickness
Theories
The Multisensory Integration Perspective (Gallagher
and Ferr
`
e, 2018) is one of the most recent frame-
works for understanding VIMS. This perspective em-
phasizes the brain’s adaptive processes and how they
might be overwhelmed in environments with diver-
gent stimuli. It proposes VIMS as the nervous sys-
tem’s challenge to appropriately weigh and integrate
various sensory signals, and it is arguably an evolu-
tion of the Neural Mismatch model (Reason, 1978b;
Oman, 1989), which is currently one of the most
widely accepted theories for VIMS. Initially proposed
for “motion sickness” alone and later applied to other
forms of VIMS, the Neural Mismatch model posits
that symptoms arise when there is a mismatch be-
tween sensory inputs, particularly between visual,
vestibular, and expectations. For example, in virtual
reality, the visual system may perceive motion, while
the vestibular system detects no movement, leading
to conflict and subsequent sickness. While this the-
ory explains many instances of VIMS, it does not
fully account for individual differences in suscepti-
bility or why some sensory mismatches cause more
severe symptoms than others, and mainly why these
would inflict symptoms.
The Postural Instability Theory (Riccio and Stof-
fregen, 1991) proposes that VIMS occurs when an
individual’s balance, which is the body’s main aim,
is disrupted as a virtual space discomfort follows.
Though some studies support this; with increased
postural sway in those experiencing sickness in vir-
tual environments, it does not explain all cases, espe-
cially in stationary, seated positions. Similarly, the
Rest Frame Hypothesis (Prothero, 1998a) suggests
the brain selects a “rest frame” or a set of stable vi-
sual references in the environment. It proposes that
sickness occurs when there are conflicting cues about
what should be considered stationary. This theory has
led to practical interventions in virtual reality, such
as adding fixed visual references to reduce sickness.
Nevertheless, all these theories are only explanations
for what triggers VIMS, and not why there would be
a trigger in the first place.
In terms of symptoms, several evolutionary ex-
planations have been proposed to explain how they
are triggered. The Poison Theory (Treisman, 1977)
suggests that the body interprets unusual sensory in-
puts as signs of poisoning, triggering nausea as a pro-
tective response. Another evolutionary perspective
proposes that sickness symptoms serve as a negative
reinforcement to discourage activities that create af-
tereffects harmful to locomotion and gaze stability
(Guedry et al., 1998). These theories offer interest-
ing perspectives on why sickness might occur; nev-
ertheless, they can struggle to explain the full range
of symptoms and individual variations and even the
counter-productivity of some strategies in critical sce-
narios.
Each of these theories contributes valuable in-
sights about VIMS, but none fully explains all aspects
of the condition. They often complement each other,
addressing different facets of the complex interplay
between human physiology and technological envi-
ronments. The limitations of these existing theories
highlight the need for a more comprehensive unifying
framework to understand and address VIMS across
various scenarios.
We propose The Exhausted Brain Theory (EBT).
A framework for understanding VIMS by integrating
concepts from most previous theories, neuroscience,
information theory, and energy metabolism. Our the-
ory posits that VIMS results from an excessive en-
ergy demand placed on the brain when it attempts to
rapidly recalibrate its internal models in response to
unfamiliar or conflicting sensory inputs.
2.2 Information Theory and
Computational Neuroscience
Information Theory, introduced by Claude Shannon
(Shannon and Weaver, 1949), provides a mathemati-
cal framework for quantifying the transmission, pro-
cessing, and storage of information, and evaluating
the efficiency of encoding schemes. It presents im-
portant concepts such as Entropy (H), Mutual Infor-
mation (I), and Channel Capacity (C), which quantify
the amount of information produced by a data source
and the reliable transmission capacity of this informa-
tion. These concepts provide the theoretical under-
pinnings for understanding information processing in
various systems, including the human brain.
The brain, as an intricate information-processing
system, can be analysed through the lens of Infor-
mation Theory. Neurons communicate via electrical
impulses, with synaptic connections facilitating the
transmission and transformation of information. Neu-
ral coding encompasses both rate coding and tempo-
ral coding, where either the firing rate of a neuron or
the timing of spikes carry information. These mecha-
nisms allow the brain to represent and process diverse
stimuli efficiently.
In sensory processing, the Efficient Coding Hy-
pothesis suggests that sensory systems are optimized
to represent information efficiently, minimizing re-
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
628

dundancy, and maximizing information transmission.
This principle is evident in phenomena such as edge
detection in the retina, which reduces redundant vi-
sual information to conserve energy and processing
capacity.
The concepts of Predictive Coding and the Free
Energy Principle (Friston, 2010), introduced by Karl
Friston, present a possible explanation of how the
brain processes information. Predictive Coding posits
that the brain continually generates predictions about
sensory inputs and updates its internal models based
on prediction errors. The Free Energy Principle ex-
tends this idea, stating that the brain seeks to mini-
mize free energy (a measure related to surprise or pre-
diction error) to maintain a stable state. These princi-
ples highlight the brain’s constant effort to balance in-
formation processing demands with energy availabil-
ity, a crucial consideration given the high metabolic
cost of neuronal activity.
2.2.1 Biological Signal Processing
The free-energy principle (Friston, 2010) proposes
that self-organizing systems maximize information
between sensory and internal states by selectively
sampling expected sensory inputs. This principle uni-
fies the Bayesian Brain Hypothesis, Efficient Cod-
ing Principle, and Cell Assembly theory, suggest-
ing the brain optimizes energy efficiency by creating
and adjusting reality models through neuronal group
changes—aligning with observed brain topology (Ma
et al., 2021).
Trujillo (Trujillo, 2019) found experimental evi-
dence that mental model adjustments increase energy
consumption and subjective exhaustion. The brain
develops multisensory integration through early-life
cross-integration training (Xu et al., 2012), enabling
mental maps crucial for spatial awareness (Hasselmo
and Stern, 2013; Hughes et al., 2014; Allen et al.,
2016). Honey et al. (Honey et al., 2012) re-
vealed complex sensory integration involving slow-
firing ”information accumulator” neurons (0.1Hz),
while Kok et al. (Kok et al., 2017) demonstrated
that expectations preactivate sensory templates in the
brain.
Neural communication is metabolically expensive
(Laughlin, 2001; Attwell and Laughlin, 2001; Lennie,
2003), consuming 35 times more energy than infor-
mation processing (Levy and Calvert, 2021). Lower
firing rates (Koch et al., 2006) and weakly active cells
(Sarpeshkar, 1998) reduce energy costs, although re-
dundancy increases the cost per bit (Laughlin, 2001).
Spatial awareness requires dense, narrow-field cells
(Sterling, 2004; Wassle and Boycott, 1991) with low
information rates to minimize costs (Koch et al.,
2006).
Brain imaging reveals that vestibular stimula-
tion reduces visual cortex blood flow (Gallagher and
Ferr
`
e, 2018; Deutschl
¨
ander et al., 2002; Wenzel et al.,
1996), while optic flow deactivates vestibular areas
(Bense et al., 2001). This apparent energy waste
(Christie and Schrater, 2015) may indicate neuronal
group decoupling for reconfiguration.
The visual system is very energy expensive; neu-
ral activity between the brain and the retina creates a
high metabolic demand. Energy is necessary for ev-
ery signal sent, and densely packed neurons are con-
stantly active (Laughlin, 2001). For mammals, 50%
of the total energy consumed by the brain is from
signaling, and in the cortex area, it represents 80%
(Laughlin, 2001).
The outputs of two adjacent photoreceptors often
measure light coming from the same object and there-
fore send very correlated signals. Thus, simply trans-
mitting their redundant information further as the out-
put of the photoreceptors would be inefficient, since
the same information would be sent multiple times
(Roland, 1999). However, in the presence of noise,
some redundancy can be helpful to (1) identify infor-
mation corruption and (2) correct errors. Thus, animal
structures such as the retina of vertebrates are made
up almost exclusively of non-spiking neurons, which
appear to be used to eliminate redundancies and noise
while boosting the remainder (Burton, 2000).
Because light varies widely in intensity and pho-
toreceptors are limited to the dynamic range, sensory
adaptation is a solution (Niven and Laughlin, 2008).
For instance, in insects, when receiving a constant in-
put, the photoreceptor will keep a tonic activity (con-
stant activation), but the neuron communication will
be phasic, allowing for the amplification and filtering
of noises. At least in insects, the output of the messag-
ing system among their neurons matches the probabil-
ity curve that maximizes information (Burton, 2000).
Moreover, humans have been observed to see signals
when pacemaker neurons are in specific phases and
miss in the opposite phase (Busch et al., 2009).
After the basics of signal reception, Field (Field,
1994) argues that natural images follow certain pat-
terns, which he describes as “sparse” (which are sim-
ilar to the filters in a CNN) and that our photorecep-
tors have arrays that activate specifically upon finding
these patterns, thus even though identifying all possi-
ble patterns requires a lot of cells, individual images
only activate a few. And then, through this method,
cognitive tasks such as learning would be facilitated
because there will be little ambiguity (Burton, 2000).
Naturally, this process can be changed because it has
been experimentally seen that the receptive fields of
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness
629
cortical cells are dynamic.
To summarize, because the visual system de-
mands a lot of energy, it must come up with ways to
be robust, effective, and energy-efficient; thus, it will
come up with representations of the world, which can
later be altered.
2.2.2 Biologically Inspired Computing
To delve deeper into the neural computations un-
derlying these principles, models such as Hopfield
Networks and Boltzmann Machines offer valuable
insights. Hopfield Networks are recurrent artificial
neural networks that function as content-addressable
memory systems. They store information in a dis-
tributed manner and retrieve it through an energy
minimization process. The network dynamics settle
into stable states (local minima of an energy func-
tion), representing stored patterns (Vallejo and Bayro-
Corrochano, 2008), (Kumar and Satsangi, 1992),
(Abubakar, 2021), (Murthy and Gabbouj, 2015).
Boltzmann Machines extend this concept by intro-
ducing stochasticity into neuron activation, allowing
the network to explore various states and escape local
minima. They are capable of learning internal rep-
resentations and modeling complex probability dis-
tributions, with an energy function (Liu and Chen,
2011), (Barra, 2012), (Agliari, 2013), (Fukai, 1992).
3 THE EXHAUSTED BRAIN
THEORY
These computational models reflect the brain’s efforts
to reach low-energy states through synaptic adjust-
ments, mirroring how neural networks adapt to mini-
mize prediction errors (Friston, 2010). In the context
of the EBT, they exemplify how processing conflict-
ing sensory inputs requires additional energy as the
brain strives to settle into a new stable state when con-
fronted with discrepancies. As such, in this case, we
will consider variations in signal that cause abnormal
activation as conflicting signals.
For example, in virtual reality or motion sim-
ulation, the usual correspondence between visual,
vestibular, and proprioceptive input is disrupted. This
mismatch increases the entropy of sensory input as
the brain faces greater uncertainty. Consequently, the
brain must process more information to resolve this
uncertainty, leading to higher entropy. This increased
entropy results in higher information processing de-
mands (Laughlin, 2001). The mutual information be-
tween sensory inputs and internal models decreases
due to the conflict, as the brain’s predictions no longer
match incoming data.
To minimize prediction errors, the brain attempts
to update its internal models, a process that requires
processing additional information and consumes sig-
nificant energy (Christie and Schrater, 2015). The
metabolic cost of these processes can lead to energy
depletion. This processing can increase entropy and
updating internal models is a metabolically demand-
ing task, with neuronal firing and synaptic plasticity
consuming energy in the form of adenosine triphos-
phate (ATP). If the energy demand exceeds supply,
the brain experiences a form of ”exhaustion,” man-
ifesting as symptoms associated with VIMS, such as
nausea and dizziness—akin to the fatigue experienced
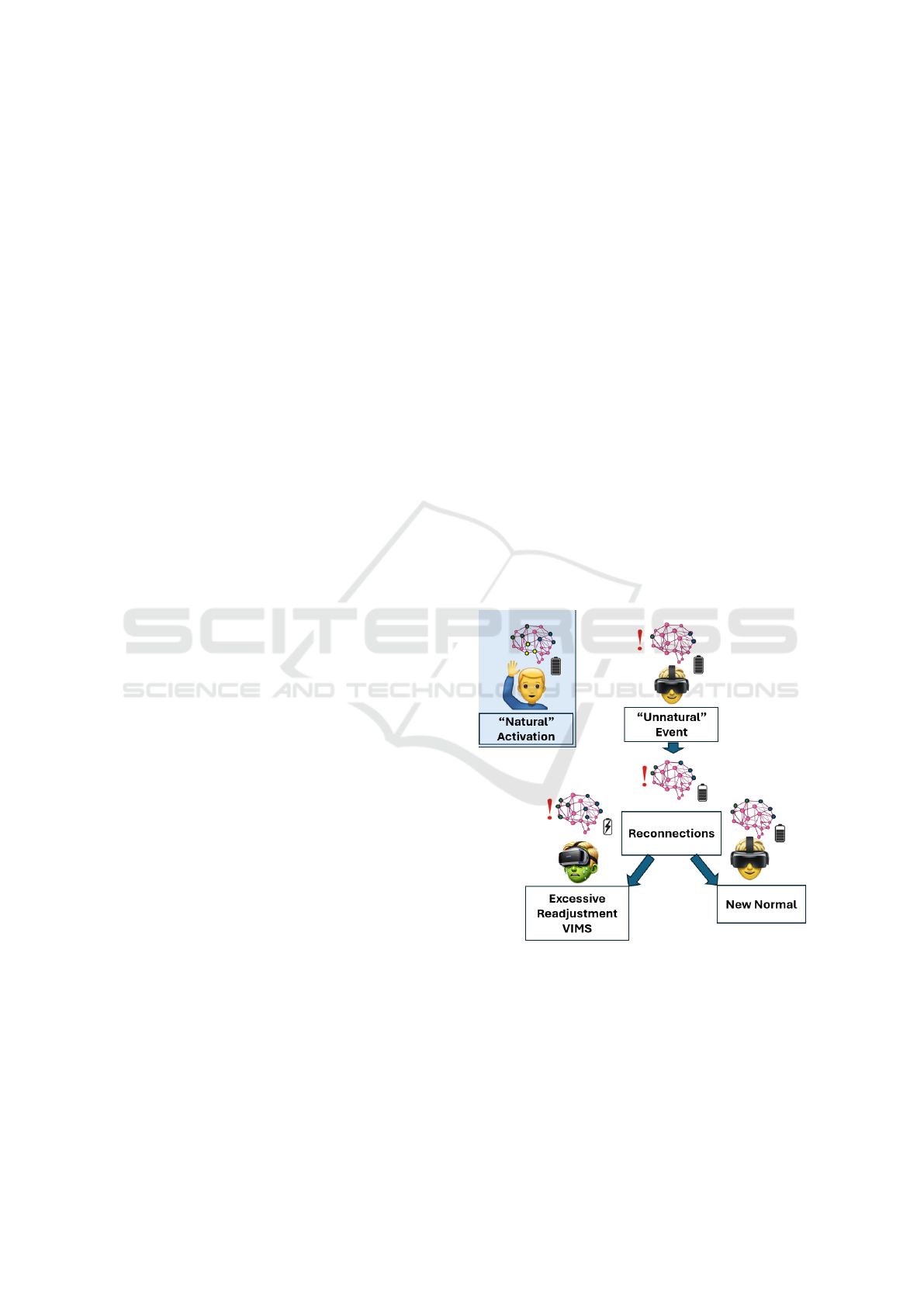
during excessive physical exercise (see Figs 1 and 2).
Channel capacity limits also play a role in VIMS.
The neural pathways have limited capacity, and ex-
cessive sensory information can overwhelm these
channels, causing delays or errors in processing. Ad-
ditionally, conflicting inputs can reduce the effective
signal-to-noise ratio, making it harder for the brain
to extract meaningful information without expending
more energy (Trujillo, 2019).
From the perspective of Predictive Coding, VIMS
can be understood as a failure of the brain’s predictive
Figure 1: A visual representation of the EBT. The user in-
teracts with an unnatural environment, such as a VR head-
set, leading to changes in the activation patterns of neurons.
This disruption requires reconnections and neural recalibra-
tion to achieve a ’new normal.’ If this recalibration occurs
efficiently, symptoms of Visually Induced Motion Sickness
(VIMS) are avoided. However, when the neural readjust-
ment is excessive or prolonged, the resulting energy deple-
tion manifests as VIMS. Neuron activations are represented
by differently colored circles.
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
630

mechanisms. Its capacity is constrained by metabolic
resources, which is why prolonged exposure to sen-
sory conflicts without adequate energy supply can
lead to persistent symptoms. The discrepancy be-
tween the expected and actual sensory inputs leads to
higher prediction errors, which are computationally
and energetically costly to resolve (Friston, 2010).
The theory also accounts for individual susceptibil-
ity to VIMS, as variations in metabolic efficiency and
neural processing capacity can affect one’s ability to
manage increased informational entropy.
Further insights come from examining the brain’s
oscillatory activity and spatial representations. Theta
rhythms, neural oscillations in the 4–8 Hz frequency
range, are prominent during active behaviors like ex-
ploration and navigation. They are associated with
memory encoding, spatial navigation, and sensorimo-
tor integration (Ravassard, 2013), (Romani, 2011),
(Zielinski et al., 2019) . Place cells, neurons in the
hippocampus, become active when an individual is in
or moving toward a specific location, forming a cog-
nitive map of the environment.
In virtual environments, sensory conflicts can dis-
rupt normal theta rhythm patterns and place cell activ-
ity, impairing the synchronization of neural networks
involved in spatial cognition. This desynchronization
requires additional neural processing to resolve, in-
creasing energy consumption. Moreover, the brain’s
effort to recalibrate its spatial maps in response to in-
consistent cues aligns with the energy demands de-
scribed in the EBT—costly, as a parallel can be drawn
from a gradient descent, in which closer values are
easier to achieve.
The concept of Neural Manifolds and Latent
Spaces provides a framework for understanding how
the brain represents high-dimensional sensory inputs
in a low-dimensional space, capturing the essential
features while reducing complexity. When sensory
inputs are conflicting or novel, this mapping becomes
less efficient, requiring more energy to process and
interpret the data. Adjusting these internal representa-
tions is metabolically demanding, contributing to the
symptoms of VIMS (Monaco, 2019), (Herweg and
Kahana, 2018), (Lu, 2020).
One of the advantages of this framework is that it
does not require the brain to have an area dedicated
to deciding when VIMS should appear and does not
impose new systems or differentiated systems to inter-
pret different inputs and detect poisoning. Moreover,
it is well accepted that “neurons that fire together wire
together”, and the ones that do not lose their connec-
tions.
This framework also highlights the importance of
energy efficiency in neural processing. It underscores
the need for virtual environments and technologies
to account for the brain’s capacity limitations and
metabolic constraints, potentially guiding the devel-
opment of interventions and design principles to mit-
igate VIMS.
In summary, the EBT synthesizes concepts from
Information Theory, computational neuroscience, and
physiological observations to explain how VIMS
arises from the brain’s overexertion due to conflicting
sensory information and the consequent energy deple-
tion. It provides a unifying framework that accounts
for individual variability and offers pathways for fu-
ture research and practical solutions.
3.1 Evidences for the Theory in
Literature
Empirical observations and research findings provide
evidence supporting the EBT’s proposition that VIMS
results from brain overexertion and energy depletion
during conflicting sensory processing. Studies have
shown that performing activities with VR headsets
leads to higher heart rates and increased calorie burn
compared to the same activities without VR, suggest-
ing increased physiological energy expenditure (Xu
et al., 2020). EEG studies indicate that task complex-
ity correlates with cybersickness (Sepich et al., 2022).
The correlation between sensory conflict and neu-
ral effort is supported by EEG research that demon-
strates higher P3 amplitudes in susceptible individuals
responding to sensory mismatches (Ahn et al., 2020).
This suggests increased cognitive demand and energy
expenditure as the brain attempts to resolve conflict-
ing sensory information. Participants who experience
symptoms also show notable changes in autonomic
responses, indicating heightened energy demand and
stress responses.
Babies and the elderly appear less susceptible to
traditional motion sickness, possibly due to differ-
ences in neural pathways or sensory reliance, ba-
bies for not having the pathways defined and the el-
derly for having a lower dependency on vestibular
cues (Schm
¨
al, 2013), thus neither needing readjust-
ment. Susceptibility factors linked to metabolic pro-
cesses further support the theory. Genetic factors are
often associated with glucose imbalance rather than
vestibular dysfunction, suggesting that efficient en-
ergy utilization is crucial to managing sensory con-
flicts (Hromatka et al., 2015).
The impact of nutrition and sleep reinforces the
EBT. Studies have shown that maintaining stable
blood sugar can mitigate symptoms, while sleep de-
privation, which impairs glucose metabolism and
cognitive function, has been linked to increased sus-
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness
631
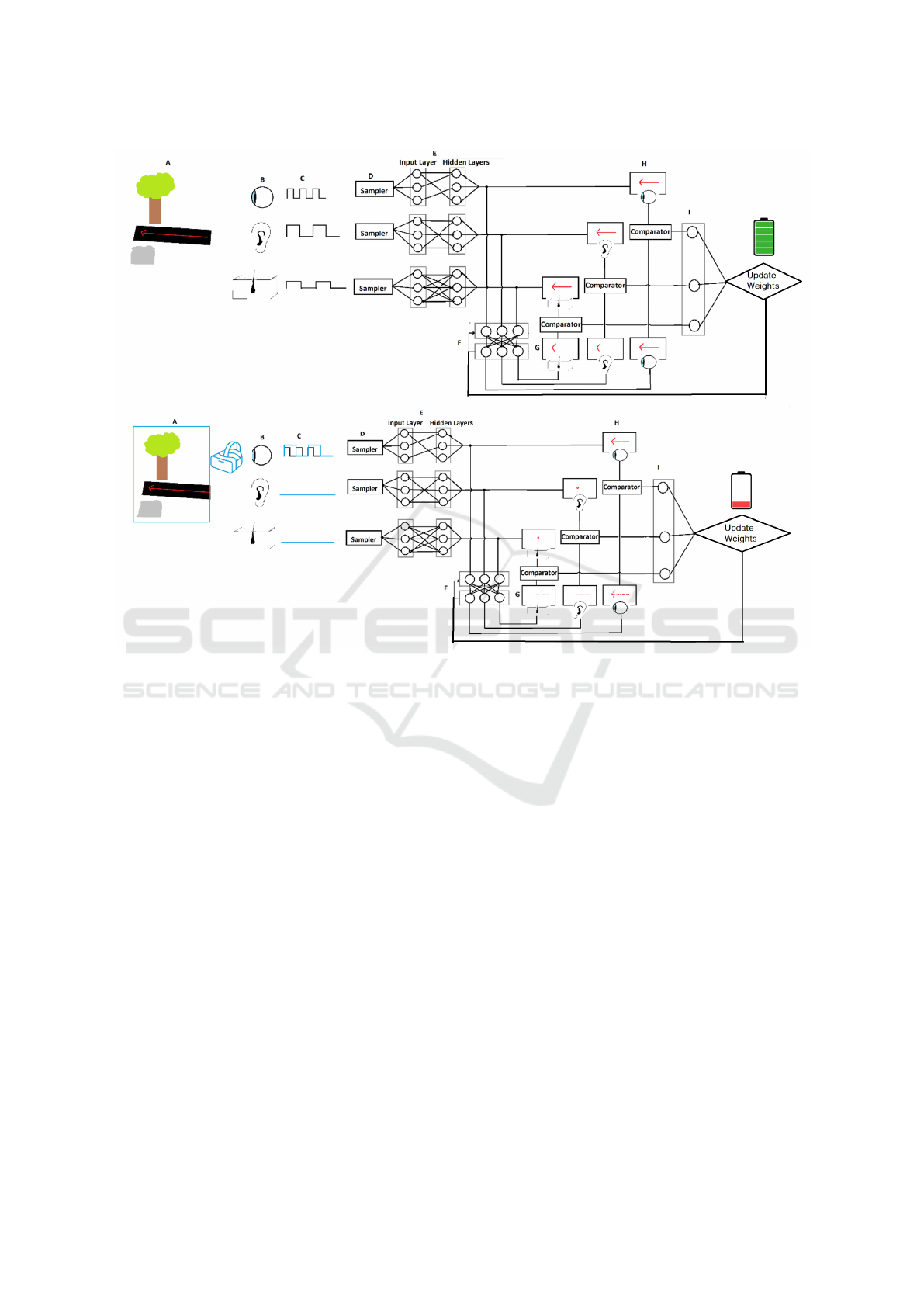
Figure 2: Top - Natural processing: Environmental signals (A) are captured by sensors (B), sampled (C,D), processed (E),
compared against predictions (G,H) based on previous states (F), and weighted (I) for continuous model updating. Bottom
- VR processing: Similar pathway but with artificial signals leading to sampling mismatches and corrupted observations,
requiring additional energy for recalibration and processing.
ceptibility (Kaplan et al., 2017). The sopite syn-
drome, characterized by drowsiness and fatigue fol-
lowing motion exposure, aligns with the theory, sug-
gesting the brain requires rest to recover and recali-
brate its energy balance (Matsangas and McCauley,
2014).
Cognitive load and learning limitations provide
further evidence. Participants using VR headsets
with higher sickness incidence showed lower rates
of knowledge acquisition, suggesting that the brain’s
resources are diverted to manage sensory conflicts
(Makransky et al., 2019). The effectiveness of miti-
gation strategies, such as gradual acclimatization and
nutritional interventions, supports the energy-based
explanation (Graybiel and Wood, 1969; Graybiel
et al., 1969).
Physiological measurements during exposure, in-
cluding altered blood flow in brain regions and elec-
trogastrographic changes, indicate metabolic activity
changes and systemic responses to neural overexer-
tion (Gavgani et al., 2018). These findings collec-
tively support the EBT, demonstrating that VIMS is
closely linked to the brain’s energy dynamics dur-
ing sensory conflicts. This comprehensive explana-
tion offers a foundation for developing targeted miti-
gation strategies focused on managing cognitive load
and supporting neural energy requirements.
3.2 Relation with Previous Theories
The EBT can encompass other theories of technol-
ogy sickness because it posits that the brain’s adap-
tation to new sensory inputs requires significant en-
ergy, leading to an ”exhausted” state when demand
exceeds supply (see table 1). The Cue Conflict The-
ory (Irwin, 1952; Bonato et al., 1990) suggests sick-
ness arises from mismatches between sensory inputs
(visual, vestibular, proprioceptive). This aligns with
the EBT because resolving these conflicts necessitates
neural rewiring, which is energy-intensive. Similarly,
the Rest Frame Hypothesis (Chang et al., 2013; Lin
et al., 2017) and Postural Instability Theory (Riccio
and Stoffregen, 1991; Villard and Flanagan, 2008) fo-
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
632

cus on the brain’s reliance on stable reference frames,
which can be integrated because recalibrating these
frames in response to conflicting cues in virtual envi-
ronments demands additional energy.
The Negative Reinforcement and Poison Theories
also fit within this framework. The former proposes
that sickness is a deterrent against potentially harm-
ful activities (Bowins, 2010), while the latter sug-
gests that it is an evolved response to perceived toxins
(Treisman, 1977; Nalivaiko et al., 2004). Both align
with the EBT because avoiding energy-depleting sit-
uations would be evolutionarily advantageous and the
residues of energy-intensive activity can be harm-
ful. Lastly, Sensory Rearrangement Theory (Reason,
1978a; Oman, 1982), which posits that the brain up-
dates paired sensory information during conflicts, and
Multisensory Integration perspectives (Gallagher and
Ferr
`
e, 2019; Kaliuzhna et al., 2015), which empha-
size the brain’s weighting of different sensory cues,
are encompassed because these processes require en-
ergy to modify neural pathways and synaptic connec-
tions. Therefore, the EBT provides a unifying frame-
work by explaining the energy demands underlying
these various theoretical perspectives.
3.3 Mitigation Through the Theory
Lenses
Several techniques can be applied to reduce VIMS,
especially in VR. For example, techniques such as the
VRCockpit (Chen et al., 2022), PlaneFrame (Mon-
teiro et al., 2020), and Rest Frames (Monteiro et al.,
2018b; Monteiro et al., 2018a; Prothero, 1998b; Shi
et al., 2021) offer consistent visual or physical cues
that help the brain establish stable reference frames,
easing the energy-intensive process of recalibration
(Wienrich et al., 2018).
Techniques that minimize sensory discrepancies
directly address the root cause of energy drain. For
example, field-of-view reduction (FOV) (Fernandes
and Feiner, 2016) and blurring (Kobayashi et al.,
2015) limit visual information, particularly in periph-
eral vision, which is the main way the brain detects
speed visually. As a result, the brain has to process
fewer conflicting cues, thereby reducing energy ex-
penditure (Lin et al., 2002).
Increasing Fidelity is an approach that can ei-
ther do wonders or have the opposite desired out-
comes. Enhancing the realism of virtual environments
by adding congruent vestibular, such as higher resolu-
tion and frame rate (Wang et al., 2023b; Wang et al.,
2022a) or proprioceptive feedback, such as vibration
or wind (Wang et al., 2022b; Zhao et al., 2024), re-
duces sensory mismatches, when done well. This can
minimize the need for the brain to reconcile discrep-
ancies, thus conserving energy (Suzuki et al., 2019).
However, presenting even more discrepant informa-
tion can exacerbate the symptoms (D’Amour et al.,
2017).
Techniques that improve the brain’s efficiency in
handling sensory input can indirectly reduce energy
consumption by optimizing information processing.
For instance, gradual exposure and adaptation by and
incremental introduction of users to virtual environ-
ments or motion stimuli allows their brain to gradu-
ally adapt and rewire neural pathways (Graybiel et al.,
1969). This staged process avoids a sudden surge in
energy demand, enabling more efficient learning and
reducing sickness over time. Even cognitive training,
with targeted exercises, can enhance the brain’s abil-
ity to process conflicting sensory information. By im-
proving efficiency, the brain can require less energy
to handle sensory discrepancies, mitigating sickness
(Nalivaiko et al., 2018).
3.4 Implications, Predictions, and
Future Work
The EBT provides a foundation for advancing the
modeling, research, treatment, detection, and mitiga-
tion of VIMS.
The brain’s energy demands during sensory con-
flicts and the resulting metabolic imbalances can be
explored using mathematical models and simulations.
These models could predict susceptibility to VIMS by
analyzing brain connectivity patterns and metabolic
rates. By focusing on specific regions and behaviors,
the accuracy of these predictions has the potential to
Table 1: Summary of what each theory accounts for. OK symbolizes that the theory accounts for that component at least
partially.
What the theories account for Sickness Triggers Individual Susceptibility Signal Intensity Why Triggers Cause Sickness Adaptation After-Effects
Cue Conflict Theory OK OK
Rest Frame Hypothesis OK OK
Postural Instability OK OK OK OK
Negative Reinforcement OK OK OK OK
Poison Theory OK OK OK OK
Sensory Rearrangement OK OK OK
Multisensory Integration OK OK OK OK OK
Exhausted Brain Theory OK OK OK OK OK OK
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness
633

improve significantly.
Furthermore, artificial neural networks could sim-
ulate VR scenarios to identify those most likely
to induce VIMS. By tracking the number of it-
erations required for the network to ”readapt” to
conflicting inputs, designers could optimize envi-
ronments to reduce VIMS triggers. For example,
a bio-inspired Spiking Neural Network trained for
self-location and mapping—using accelerometer and
video data—could later be exposed to new VIMS-like
”noisy” data. The time required for readaptation in re-
sponse to this interference could serve as a guideline
for detecting VIMS-triggering environments.
The development of a standard for movement pa-
rameters and dimensions in XR environments could
also facilitate broader adoption. Mental training
acquired in one environment might transfer to an-
other application. However, applications with un-
even movement patterns or inconsistent frame opti-
mization could still cause users to struggle. For in-
stance, in a single application, fluctuating frame rates
are more likely to induce sickness. This theory high-
lights that individual differences in sensory integra-
tion and metabolic efficiency influence susceptibility
to VIMS. It also underscores that adaptation to one
type of VIMS-inducing environment may not guar-
antee immunity in others, emphasizing the need for
tailored acclimatization.
Given this context, we can expect users who
present better spatial acuity and use several cues for
self-location to suffer more from VIMS than those
with poor self-location.
Using EEG to synchronize refresh rates with neu-
ronal activation frequencies—-potentially even vary-
ing by screen area—-may reduce VIMS. This ap-
proach could allow for better resource allocation, as
not all screen regions may need simultaneous updates.
Developing specific tools to measure energy ex-
penditure and adaptation processes could improve
VIMS detection. For example, functional near-
infrared spectroscopy (fNIRS) or similar non-invasive
methods could help analyze neural and metabolic re-
sponses. Recording blood glucose levels before and
after VR exposure, where ethically permissible, could
reveal metabolic changes linked to VIMS. Partici-
pants could also be asked about conditions affecting
glucose processing, such as diabetes, pancreatic dis-
orders, or body mass, to refine the collected data.
Given the nature of the theory, it would theoret-
ically be possible to assess an individual’s suscepti-
bility by analyzing information beyond the exposure
itself. This could involve determining the strength of
the coupling between their visual and vestibular sys-
tems, as well as evaluating the amount of energy re-
quired to readjust when faced with a different form of
conflict. In a longitudinal study, as the degradation
of the vestibular system slowly causes the decoupling
of the two information systems, VIMS should be less
present (considering all other metabolic aspects re-
main constant).
For long-term studies or repeated exposure, intro-
ducing a ”Day-0” adaptation protocol for users and
participants would be ideal. This protocol would al-
low individuals to acclimate to new simulators and
HMDs, ensuring that they experience applications
as intended. Structured acclimatization frameworks
might include 1) Exercises to ”decouple” visual and
vestibular senses. 2) Gradual exposure to VIMS-
inducing content, starting with simpler environments
(e.g., figure outlines) and increasing complexity in-
crementally. 3) Training strategies to optimize brain
energy usage, akin to stepwise progression in athletic
training. 4) Adequate nutritional intake.
Detecting when users are ready to engage in learn-
ing activities within VIMS-inducing environments
could enhance the effectiveness of these settings. En-
suring habituation before introducing cognitive tasks
could prevent overload and maximize learning out-
comes. Short-term studies should consider using par-
ticipants who are already acclimated to the environ-
ment to avoid skewing the results. Moreover, studies
showing minimal learning differences in VR may un-
derestimate its potential, as new exposure to VR could
hinder outcomes. This suggests that some findings
that consider VR ineffective for learning might stem
from studies that are too short (Monteiro et al., 2024;
Barrett et al., 2023; Makransky et al., 2019).
The EBT could also inform new health and safety
guidelines for prolonged XR and simulator use, par-
ticularly in professional contexts where extended use
is required. Additionally, it raises important con-
siderations for children’s use of XR and simulators.
As ”virtual natives,” children exposed to XR envi-
ronments may develop long-term advantages, such
as more adaptable movement pathways and increased
neural connections, akin to the benefits of early lan-
guage acquisition, once thought to be a disadvantage.
This also means that it is possible that some people
could never adapt when exposed to just ”normal” use.
3.5 Testable Hypotheses
To validate the EBT, several testable hypotheses can
be proposed. These hypotheses aim to explore the
relationship between energy dynamics, sensory con-
flicts, and individual variability in the context of Vi-
sually Induced Motion Sickness.
H1: Individuals performing cognitively demand-
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
634

ing tasks during exposure to a sensory conflict-
inducing VR environment will experience higher lev-
els of VIMS compared to those in less demanding
tasks. A controlled experiment could compare VIMS
to physiological markers of energy expenditure.
H2: Gradual acclimatization to VR environments
will reduce VIMS symptoms over time compared to
abrupt exposure to complex environments. This hy-
pothesis could be explored in a longitudinal study by
comparing symptoms between participants exposed
to incremental versus sudden increases in the com-
plexity of the VR session.
H3: Individuals with a higher baseline efficiency
in predictive coding will experience less severe VIMS
in sensory-conflicting VR environments. Predictive
coding efficiency could be measured using EEG pat-
terns or behavioral tests of sensory integration and
correlated with VIMS symptoms.
H4: Increasing the sensory fidelity of VR environ-
ments, such as by adding congruent vestibular feed-
back in an extremely precise way, will reduce VIMS
symptoms compared to conditions with sensory mis-
matches; otherwise, it will cause worse symptoms.
This could be tested by comparing symptoms and
EEG markers such as theta rhythm desynchronization
in conditions with and without synchronized vestibu-
lar feedback.
H5: Participants with higher aerobic fitness lev-
els will exhibit lower VIMS susceptibility, as better
metabolic efficiency could buffer against the energy
depletion associated with sensory conflicts.
H6: Participants with more stable theta rhythms
during VR exposure will experience fewer VIMS
symptoms. EEG data could be used to examine theta
rhythm stability and its relationship with VIMS sever-
ity across varying sensory conflict levels.
These hypotheses provide a framework for empir-
ical validation of the EBT and should be tested us-
ing techniques that detect VIMS within the experi-
ment (Wang et al., 2020; Wang et al., 2023a; Mon-
teiro et al., 2021).
3.6 Limitations
One limitation of this work is its correlational nature,
as it does not include empirical evidence derived from
original experiments. While correlations cannot es-
tablish causation, this does not diminish the valid-
ity of the framework presented. Similar research in
this field also relies on the existing literature and ob-
servational studies, and other recognized theories are
untestable. Furthermore, the correlations drawn here
are supported by multiple reliable sources, lending
credibility to the insights. Importantly, this work is
intended as a foundation for future research, provid-
ing a framework that can guide experiments designed
to either support or challenge this theory.
Another limitation of this paper is that we do not
delve into the specifics of how glucose is processed
or explore the broader metabolic processes in detail.
Consequently, the root cause of the issue might be
related to the energy demand process rather than the
energy demand itself. However, this lies beyond the
scope of the paper and the technical expertise of the
researchers. Despite this limitation, related studies
still support the overall association between energy
and VIMS. The authors remain open to alternative
interpretations, e.g., such as the possibility that the
problem might stem from residual effects of the read-
justment process rather than the readjustment itself.
4 CONCLUSION
The EBT provides a comprehensive and unifying
framework to understand the phenomenon of VIMS.
By emphasizing the brain’s metabolic demands and
energy limitations during sensory conflict resolution,
the theory integrates insights from established per-
spectives, such as sensory conflict, postural instabil-
ity, and multisensory integration. It highlights how
the brain’s effort to recalibrate internal models in re-
sponse to conflicting sensory inputs can lead to energy
depletion and, consequently, VIMS symptoms.
This energy-based approach not only explains in-
dividual susceptibilities to VIMS but also accounts
for the variability in symptoms across scenarios and
technologies. By framing VIMS as a product of the
brain’s effort to minimize prediction errors and main-
tain internal stability, the theory bridges the gap be-
tween neurophysiology, computational neuroscience,
and energy metabolism. It underscores the impor-
tance of designing energy-efficient virtual environ-
ments and personalized interventions to reduce the
cognitive and metabolic strain associated with immer-
sive technologies.
Furthermore, the EBT lays the groundwork for fu-
ture research aimed at modeling, detecting, and miti-
gating VIMS. From developing tools to measure neu-
ral and metabolic responses to designing acclimatiza-
tion protocols and energy-aware XR systems, the the-
ory inspires practical solutions to a critical challenge
in the adoption of emerging technologies. As immer-
sive environments become increasingly integral to ed-
ucation, healthcare, and entertainment, understanding
and addressing the underlying causes of VIMS is es-
sential for their safe and effective use.
In conclusion, the EBT not only enriches our un-
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness
635

derstanding of VIMS but also opens new avenues for
research and innovation, offering a pathway toward
enhancing the accessibility and usability of virtual
and augmented reality technologies.
REFERENCES
Abubakar, H. (2021). An optimal representation of random
maximum ksatisfiability on a hopfield neural network
for high order logic (k ¿ 3). Kuwait Journal of Science.
Agliari, E. e. a. (2013). Parallel retrieval of correlated
patterns: From hopfield networks to boltzmann ma-
chines. Neural Networks.
Ahn, M.-H., Park, J., Jeon, H., Lee, H.-J., Kim, H.-J., and
Hong, S. (2020). Temporal dynamics of visually in-
duced motion perception and neural evidence of alter-
ations in the motion perception process in an immer-
sive virtual reality environment. Frontiers in Neuro-
science, 14:600839. Received: 31 August 2020; Ac-
cepted: 29 October 2020; Published: 19 November
2020.
Allen, B., Hanley, T., Rokers, B., and Green, C. S. (2016).
Visual 3d motion acuity predicts discomfort in 3d
stereoscopic environments. Entertainment Comput-
ing, 13:1–9.
Attwell, D. and Laughlin, S. B. (2001). An energy budget
for signaling in the grey matter of the brain. Journal
of Cerebral Blood Flow & Metabolism.
Barra, A. e. a. (2012). On the equivalence of hopfield net-
works and boltzmann machines. Neural Networks.
Barrett, A., Pack, A., Monteiro, D., and Liang, H.-N.
(2023). Exploring the influence of audience famil-
iarity on speaker anxiety and performance in virtual
reality and real-life presentation contexts. Behaviour
& Information Technology, 43(4):787–799.
Bense, S., Stephan, T., Yousry, T. A., Brandt, T., and
Dieterich, M. (2001). Multisensory cortical sig-
nal increases and decreases during vestibular gal-
vanic stimulation (fmri). Journal of neurophysiology,
85(2):886–899.
Bonato, F., Bubka, A., and Palmisano, S. (1990). Sensory
conflict and simulator sickness: An experiment with
multi-axis motion. Aviation, Space, and Environmen-
tal Medicine, 61:690–693.
Bowins, B. (2010). Motion sickness: A defense mecha-
nism? Medical Hypotheses, 74:661–663.
Burton, G. G. (2000). Problems and solutions in early vi-
sual processing. In Information Theory and The Brain.
Cambridge University Press.
Busch, N. A., Dubois, J., and VanRullen, R. (2009). The
phase of ongoing eeg oscillations predicts visual per-
ception. The Journal of Neuroscience, 29(24):7869–
7876.
Chang, E., Kim, T., and Yoo, S. (2013). Reducing cy-
bersickness using a visual grid. In Proceedings of
the ACM Symposium on Virtual Reality Software and
Technology (VRST), pages 99–106.
Chen, H., Shi, R., Monteiro, D., Baghaei, N., and Liang, H.-
N. (2022). Vrcockpit: Mitigating simulator sickness
in vr games using multiple egocentric 2d view frames.
In 2022 IEEE Conference on Games (CoG), pages 80–
87.
Christie, S. T. and Schrater, P. (2015). Cognitive cost as
dynamic allocation of energetic resources. Frontiers
in neuroscience, 9:289.
D’Amour, S., Bos, J. E., and Keshavarz, B. (2017). The
efficacy of airflow and seat vibration on reducing vi-
sually induced motion sickness. Experimental Brain
Research, 235:2811–2820.
Deutschl
¨
ander, A., Bense, S., Stephan, T., Schwaiger, M.,
Brandt, T., and Dieterich, M. (2002). Sensory system
interactions during simultaneous vestibular and visual
stimulation in pet. Human Brain Mapping, 16(2):92–
103.
Dobie, T. G. (2019). Motion Sickness: A Motion Adaptation
Syndrome. Springer.
Fernandes, A. S. and Feiner, S. K. (2016). Combating vr
sickness through subtle dynamic field-of-view modi-
fication. In IEEE Symposium on 3D User Interfaces.
Field, D. J. (1994). What is the goal of sensory coding?
Neural Computation, 6(4):559–601.
Friston, K. (2010). The free-energy principle: A uni-
fied brain theory? Nature Reviews Neuroscience,
11(2):127–138.
Fukai, T. (1992). Comparative study of spurious-state dis-
tribution in analogue neural networks and boltzmann
machine. Journal of Physics A.
Gallagher, M. and Ferr
`
e, E. R. (2018). Cybersickness:
A multisensory integration perspective. Multisensory
Research, 31(7):645–674.
Gallagher, M. and Ferr
`
e, E. (2019). The multisensory in-
tegration perspective: A framework for understanding
motion sickness. Neuroscience Letters, 706:33–39.
Gavgani, A. M., Wong, R. H., Howe, P. R., Hodgson,
D. M., Walker, F. R., and Nalivaiko, E. (2018).
Cybersickness-related changes in brain hemodynam-
ics: A pilot study comparing transcranial doppler and
near-infrared spectroscopy assessments during a vir-
tual ride on a roller coaster. Physiology & behavior,
191:56–64.
Graybiel, A., Deane, F. R., and Colehour, J. K. (1969). Pre-
vention of overt motion sickness by incremental ex-
posure to otherwise highly stressful coriolis accelera-
tions. Aerospace medicine, 40(2):142–148.
Graybiel, A. and Wood, C. D. (1969). Rapid vestibu-
lar adaptation in a rotating environment by means
of controlled head movements. Aerospace medicine,
40(6):638–643.
Guedry, F. E., Rupert, A. H., and Reschke, M. F. (1998).
Motion sickness and development of synergy within
the spatial orientation system. a hypothetical unifying
concept. Brain Research Bulletin, 47(5):475–480.
Hasselmo, M. E. and Stern, C. E. (2013). Theta rhythm and
the encoding and retrieval of space and time. Neu-
roImage.
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
636

Herweg, N. A. and Kahana, M. (2018). Spatial representa-
tions in the human brain. Frontiers in Human Neuro-
science.
Honey, C. J., Thesen, T., Donner, T. H., Silbert, L. J., Carl-
son, C. E., Devinsky, O., Doyle, W. K., Rubin, N.,
Heeger, D. J., and Hasson, U. (2012). Slow cortical
dynamics and the accumulation of information over
long timescales. Neuron, 76(2):423–434.
Hromatka, B. S., Tung, J. Y., Kiefer, A. K., Do, C. B.,
Hinds, D. A., and Eriksson, N. (2015). Genetic vari-
ants associated with motion sickness point to roles
for inner ear development, neurological processes and
glucose homeostasis. Human Molecular Genetics,
24(9):2700–2708.
Hughes, M., Sulikowski, D., and Burke, D. (2014). Cor-
relations between spatial skills: A test of the hunter-
gatherer hypothesis. Journal of Evolutionary Psychol-
ogy, 12(1):19–44.
Irwin, J. (1952). The pathology of seasickness. The Lancet,
2:907–909.
Kaliuzhna, M., Prsa, M., and Blanke, O. (2015). Vestibu-
lar reweighting and multisensory integration in cyber-
sickness. PLoS ONE, 10:e0138563.
Kaplan, J., Ventura, J., Bakshi, A., Pierobon, A., Lackner,
J. R., and DiZio, P. (2017). The influence of sleep
deprivation and oscillating motion on sleepiness, mo-
tion sickness, and cognitive and motor performance.
Autonomic Neuroscience, 202:86–96.
Kobayashi, N., Iinuma, R., Suzuki, Y., Shimada, T., and
Ishikawa, M. (2015). Using bio-signals to evaluate
multi-discomfort in image viewing: Balancing visu-
ally induced motion sickness and field of view. In
37th Annual International Conference of the IEEE En-
gineering in Medicine and Biology Society (EMBC).
Koch, K. et al. (2006). How much the eye tells the brain.
Current Biology.
Kok, P., Mostert, P., and de Lange, F. P. (2017). Prior
expectations induce prestimulus sensory templates.
Proceedings of the National Academy of Sciences,
114(39):10473–10478.
Kumar, S. and Satsangi, P. (1992). System dynamics simu-
lation of hopfield neural networks. International Jour-
nal of Systems Science.
Laughlin, S. B. (2001). Energy as a constraint on the cod-
ing and processing of sensory information. Current
Opinion in Neurobiology, 11(4):475–480.
Lennie, P. (2003). The cost of cortical computation. Current
Biology.
Levy, W. B. and Calvert, V. G. (2021). Communication
consumes 35 times more energy than computation in
the human cortex.
Lin, J., Duh, H., and Parker, D. (2017). Effects of rest
frames on cybersickness in a virtual environment.
IEEE Transactions on Visualization and Computer
Graphics, 23(4):1255–1264.
Lin, J. J., Abi-Rached, H., Kim, D. H., Parker, D. E., and
Furness, T. A. (2002). A ’natural’ independent visual
background reduced simulator sickness. In Proceed-
ings of the Human Factors and Ergonomics Society
Annual Meeting.
Liu, C.-C. and Chen, C. (2011). Set based boltzmann ma-
chine and hopfield neural networks. NANO.
Lu, Y. (2020). Hippocampal spatial representation: Inte-
grating environmental and self-motion signals. Fron-
tiers in Cellular Neuroscience.
Ma, J., Zhang, Y., Lin, Y., and Dai, Z. (2021). Cost-
efficiency trade-offs of the human brain network re-
vealed by a multiobjective evolutionary algorithm.
Neuroimage, 236.
Makransky, G., Terkildsen, T. S., and Mayer, R. E. (2019).
Adding immersive virtual reality to a science lab sim-
ulation causes more presence but less learning. Learn-
ing and Instruction, 60:225–236.
Matsangas, P. and McCauley, M. E. (2014). Yawning as
a behavioral marker of mild motion sickness and so-
pite syndrome. Aviation, Space, and Environmental
Medicine, 85(6).
Monaco, J. e. a. (2019). Spatial synchronization codes from
coupled rate-phase neurons. PLOS Computational Bi-
ology.
Monteiro, D., Liang, H.-N., Abel, A., Baghaei, N., and
Monteiro, R. d. C. (2018a). Evaluating engagement
of virtual reality games based on first and third per-
son perspective using eeg and subjective metrics. In
2018 IEEE International Conference on Artificial In-
telligence and Virtual Reality (AIVR), pages 53–60.
Monteiro, D., Liang, H.-N., Tang, X., and Irani, P. (2021).
Using trajectory compression rate to predict changes
in cybersickness in virtual reality games. In 2021
IEEE International Symposium on Mixed and Aug-
mented Reality (ISMAR), pages 138–146.
Monteiro, D., Liang, H.-N., Wang, J., Chen, H., and
Baghaei, N. (2020). An in-depth exploration of the ef-
fect of 2d/3d views and controller types on first person
shooter games in virtual reality. In 2020 IEEE Inter-
national Symposium on Mixed and Augmented Reality
(ISMAR), pages 713–724.
Monteiro, D., Liang, H.-N., Zhao, Y., and Abel, A. (2018b).
Comparing event related arousal-valence and focus
among different viewing perspectives in vr gaming. In
Advances in Brain Inspired Cognitive Systems. BICS
2018. Lecture Notes in Computer Science, volume
10989. Springer, Cham.
Monteiro, D., Wang, A., Wang, L., et al. (2024). Effects
of audience familiarity on anxiety in a virtual reality
public speaking training tool. Universal Access in the
Information Society, 23:23–34.
Murthy, G. R. and Gabbouj, M. (2015). On the design of
hopfield neural networks: Synthesis of hopfield type
associative memories. IJCNN.
Nalivaiko, E., Gavgani, A. M., and Walker, F. R. (2018).
Cybersickness-related changes in brain hemodynam-
ics: A pilot study comparing transcranial doppler and
near-infrared spectroscopy assessments during a vir-
tual ride on a roller coaster. Physiology & Behavior,
191:56–64.
Nalivaiko, E., Rudd, J., and So, R. (2004). Motion sick-
ness, nausea, and temperature changes: A hypothesis.
Autonomic Neuroscience, 116:77–82.
The Exhausted Brain Theory: An Energy-Based Framework for Understanding Visually Induced Motion Sickness
637

Niven, J. E. and Laughlin, S. B. (2008). Energy lim-
itation as a selective pressure on the evolution of
sensory systems. Journal of Experimental Biology,
211(11):1792–1804.
Oman, C. (1982). A heuristic mathematical model for
the dynamics of sensory conflict and motion sickness.
Acta Oto-Laryngologica Supplementum, 392:1–44.
Oman, C. M. (1989). Sensory conflict in motion sickness:
An observer theory approach.
Prothero, J. D. (1998a). The role of rest frames in vection,
presence and motion.
Prothero, J. D. (1998b). The role of rest frames in vection,
presence and motion. In Proceedings of the IEEE Vir-
tual Reality Conference.
Ravassard, P. e. a. (2013). Multisensory control of hip-
pocampal spatiotemporal selectivity. Science.
Reason, J. (1978a). Motion sickness adaptation: A neu-
ral mismatch model. Journal of the Royal Society of
Medicine, 71:819–829.
Reason, J. (1978b). Motion sickness: Some theoretical
and practical considerations. Applied Ergonomics,
9(3):163–167.
Riccio, G. E. and Stoffregen, T. (1991). An ecological the-
ory of motion sickness and postural instability.
Roland, B. (1999). Introductory information theory and the
brain.
Romani, S. e. a. (2011). Intracellular dynamics of virtual
place cells. Neural Computation.
Sarpeshkar, R. (1998). Analog versus digital: extrapolating
from electronics to neurobiology. Neural Computa-
tion.
Schm
¨
al, F. (2013). Neuronal mechanisms and the treatment
of motion sickness. Pharmacology, 91(3-4):229–241.
Sepich, N. C., Jasper, A., Fieffer, S., Gilbert, S. B., Dorne-
ich, M. C., and Kelly, J. W. (2022). The impact of
task workload on cybersickness. Frontiers in Virtual
Reality, 0:110.
Shannon, C. E. and Weaver, W. (1949). The Mathemati-
cal Theory of Communication. University of illinois
Press.
Shi, R., Liang, H.-N., Wu, Y., Yu, D., and Xu, W. (2021).
Virtual reality sickness mitigation methods: A com-
parative study in a racing game. Proc. ACM Comput.
Graph. Interact. Tech., 4(1).
Sterling, P. (2004). How retinal circuits optimize the trans-
fer of visual information.
Suzuki, Y., Yem, V., Hirota, K., Amemiya, T., Kitazaki, M.,
and Ikei, Y. (2019). Airflow presentation method for
turning motion feedback in vr environment. In ICAT-
EGVE 2019 - International Conference on Artificial
Reality and Telexistence Eurographics Symposium on
Virtual Environments.
Treisman, M. (1977). Motion sickness: An evolutionary
hypothesis. Science, 197(4302):493–495.
Trujillo, L. T. (2019). Mental effort and information-
processing costs are inversely related to global brain
free energy during visual categorization. Frontiers in
Neuroscience, 13.
Vallejo, R. and Bayro-Corrochano, E. (2008). Clifford hop-
field neural networks. IJCNN.
Villard, S. and Flanagan, P. (2008). Postural activity pre-
dicts simulator sickness. Presence: Teleoperators and
Virtual Environments, 17:517–532.
Wang, J., Liang, H.-N., Monteiro, D., Xu, W., and Xiao, J.
(2023a). Real-time prediction of simulator sickness in
virtual reality games. IEEE Transactions on Games,
15(2):252–261.
Wang, J., Liang, H.-N., Monteiro, D. V., Xu, W., Chen, H.,
and Chen, Q. (2020). Real-time detection of simula-
tor sickness in virtual reality games based on players’
psychophysiological data during gameplay. In 2020
IEEE International Symposium on Mixed and Aug-
mented Reality Adjunct (ISMAR-Adjunct), pages 247–
248.
Wang, J., Shi, R., Xiao, Z., Qin, X., and Liang, H.-N.
(2022a). Effect of render resolution on gameplay ex-
perience, performance, and simulator sickness in vir-
tual reality games. Proc. ACM Comput. Graph. Inter-
act. Tech., 5(1).
Wang, J., Shi, R., Zheng, W., Xie, W., Kao, D., and Liang,
H.-N. (2023b). Effect of frame rate on user experi-
ence, performance, and simulator sickness in virtual
reality. IEEE Transactions on Visualization and Com-
puter Graphics, 29(5):2478–2488.
Wang, X., Monteiro, D., Lee, L.-H., Hui, P., and Liang, H.-
N. (2022b). Vibroweight: Simulating weight and cen-
ter of gravity changes of objects in virtual reality for
enhanced realism. In 2022 IEEE Haptics Symposium
(HAPTICS), pages 1–7.
Wassle, H. and Boycott, B. (1991). Functional architecture
of the mammalian retina. Physiological Reviews.
Wenzel, R., Bartenstein, P., Dieterich, M., Danek, A.,
Weindl, A., Minoshima, S., Ziegler, S., Schwaiger,
M., and Brandt, T. (1996). Deactivation of human vi-
sual cortex during involuntary ocular oscillations: A
pet activation study. Brain, 119(1):101–110.
Wienrich, C., Weidner, C. K., Schatto, C., Obremski, D.,
and Israel, J. H. (2018). A virtual nose as a rest frame:
The impact on simulator sickness and game experi-
ence. In 10th International Conference on Virtual
Worlds and Games for Serious Applications.
Xu, J., Yu, L., Rowland, B. A., Stanford, T. R., and Stein,
B. E. (2012). Incorporating cross-modal statistics in
the development and maintenance of multisensory in-
tegration. Journal of Neuroscience, 32(7):2287–2298.
Xu, W., Liang, H.-N., Zhang, Z., and Baghaei, N. (2020).
Studying the effect of display type and viewing per-
spective on user experience in virtual reality ex-
ergames. Games for Health Journal, 9(4):1–10.
Zhao, F., Li, Z., Luo, Y., Li, Y., and Liang, H.-N. (2024).
AirWhisper: enhancing virtual reality experience via
visual-airflow multimodal feedback. Journal on Mul-
timodal User Interfaces.
Zielinski, M. C., Shin, J. D., and Jadhav, S. P. (2019). Co-
herent coding of spatial position mediated by theta os-
cillations in the hippocampus and prefrontal cortex.
Journal of Neuroscience, 39(20):4026–4040.
HUCAPP 2025 - 9th International Conference on Human Computer Interaction Theory and Applications
638
